Прочетете биохимията на мускулната активност на вълците. Динамика на биохимичните процеси в организма по време на мускулна работа. контракции, общ преглед на скелетните мускули)
Изпратете добрата си работа в базата знания е лесно. Използвайте формата по-долу
Студенти, докторанти, млади учени, които използват базата от знания в обучението и работата си, ще ви бъдат много благодарни.
публикувано на http://www.allbest.ru/
Въведение
1. Скелетни мускули, мускулни протеини и биохимични процеси в мускулите
2. Биохимични промени в тялото на спортистите по бойни изкуства
4. Проблемът за възстановяването в спорта
5. Характеристики на метаболитните състояния при хора по време на мускулна дейност
6. Биохимичен контрол в бойните изкуства
Заключение
Библиография
Въведение
Ролята на биохимията в съвременната спортна практика нараства все повече. Без познаване на биохимията на мускулната активност и механизмите на метаболитна регулация по време на физически упражнения е невъзможно ефективното управление на тренировъчния процес и неговата по-нататъшна рационализация. Познаването на биохимията е необходимо за оценка на нивото на годност на спортиста, идентифициране на претоварвания и пренапрежение, както и за правилното организиране на диета. Една от най-важните задачи на биохимията е да намери ефективни начини за контролиране на метаболизма въз основа на дълбоки познания за химичните трансформации, тъй като състоянието на метаболизма определя нормалността и патологията. Растежът и развитието на живия организъм, способността му да издържа на външни влияния и активно да се адаптира към новите условия на съществуване зависи от естеството и скоростта на метаболитните процеси.
Изследването на адаптивните промени в метаболизма ни позволява да разберем по-добре характеристиките на адаптацията на тялото към физическа активност и да намерим ефективни средства и методи за повишаване на физическата работоспособност.
В бойните спортове проблемът с физическата годност винаги се е считал за един от най-важните, определящи нивото на спортните постижения.
Обичайният подход за определяне на тренировъчните методи се основава на емпирични закони, които формално описват явленията на спортната тренировка.
Самите физически качества обаче не могат да съществуват сами по себе си. Те се появяват в резултат на това, че централната нервна система контролира мускулите, които се свиват и губят метаболитна енергия.
Теоретичният подход изисква изграждането на модел на тялото на спортиста, като се вземат предвид постиженията на световната спортна биология. За да се контролират процесите на адаптация в определени клетки на органите на човешкото тяло, е необходимо да се знае структурата на органа, механизмите на неговото функциониране и факторите, които осигуряват целевата посока на процесите на адаптация.
1. Скелетни мускули, мускулни протеини и биохимични процеси в мускулите
Скелетните мускули съдържат голямо количество непротеинови вещества, които лесно преминават от смачкани мускули във воден разтвор след утаяване на протеини. АТФ е пряк източник на енергия не само за различни физиологични функции (мускулни контракции, нервна дейност, предаване на нервно възбуждане, процеси на секреция и др.), Но също така и за пластични процеси, протичащи в тялото (изграждане и обновяване на тъканни протеини, биологични синтези). Съществува постоянна конкуренция между тези две страни на живота - енергийното снабдяване на физиологичните функции и енергийното снабдяване на пластичните процеси. Изключително трудно е да се дадат определени стандартни норми за биохимичните промени, които настъпват в тялото на спортиста при практикуване на един или друг спорт. Дори при извършване на отделни упражнения в чист вид (лекоатлетическо бягане, кънки, ски), протичането на метаболитните процеси може да се различава значително при различните спортисти в зависимост от вида на нервната им дейност, влиянието на околната среда и др. Скелетната мускулатура съдържа 75-80% вода и 20-25% сухо вещество. 85% от сухия остатък са протеини; останалите 15% се състоят от различни азотсъдържащи и безазотни екстракти, фосфорни съединения, липоиди и минерални соли. Мускулни протеини. Саркоплазмените протеини съставляват до 30% от всички мускулни протеини.
Мускулните фибрилни протеини съставляват около 40% от всички мускулни протеини. Протеините на мускулните фибрили включват предимно два основни протеина - миозин и актин. Миозинът е протеин от глобулинов тип с молекулно тегло около 420 000. Съдържа много глутаминова киселина, лизин и левцин. В допълнение, заедно с други аминокиселини, той съдържа цистеин и следователно има свободни групи - SH. Миозинът е разположен в мускулните фибрили в дебели нишки на "диск А" и не хаотично, а строго подреден. Молекулите на миозина имат нишковидна (фибриларна) структура. Според Хъксли тяхната дължина е около 1500 А, дебелината е около 20 А. Имат удебеление в единия край (40 А). Тези краища на неговите молекули са насочени в двете посоки от "М зоната" и образуват клубообразни удебеления на процесите на дебели нишки. Миозинът е основен компонент на контрактилния комплекс и в същото време има ензимна (аденозин трифосфатазна) активност, като катализира разграждането на аденозин трифосфорната киселина (АТФ) до ADP и ортофосфат. Актинът има много по-малко молекулно тегло от миозина (75 000) и може да съществува в две форми – глобуларна (G-актин) и фибриларна (F-актин), способни да се трансформират една в друга. Молекулите на първия имат кръгла форма; втората молекула, която е полимер (комбинация от няколко молекули) на G-актин, е нишковидна. G-актинът има нисък вискозитет, F-актинът има висок вискозитет. Преходът на една форма на актин в друга се улеснява от много йони, по-специално K+ и Mg++. По време на мускулна активност G-актинът се трансформира във F-актин. Последният лесно се свързва с миозина, образувайки комплекс, наречен актомиозин, и е контрактилен субстрат на мускула, способен да произвежда механична работа. В мускулните фибрили актинът се намира в тънки нишки на "J диска", простирайки се в горната и долната третина на "А диска", където актинът е свързан с миозина чрез контакти между процесите на тънки и дебели нишки. В допълнение към миозина и актина, някои други протеини също са открити в миофибрилите, по-специално водоразтворимият протеин тропомиозин, който е особено изобилен в гладките мускули и в мускулите на ембрионите. Фибрилите съдържат и други водоразтворими протеини, които имат ензимна активност” (деаминаза на аденилова киселина и др.). Протеините на митохондриите и рибозомите са главно ензимни протеини. По-специално, митохондриите съдържат ензими на аеробно окисление и респираторно фосфорилиране, а рибозомите съдържат свързана с протеин rRNA. Протеините на ядрата на мускулните влакна са нуклеопротеини, съдържащи дезоксирибонуклеинови киселини в техните молекули.
Протеини на стромата на мускулните влакна, съставляващи около 20% от всички мускулни протеини. От стромални протеини, наречени от A.Ya. Данилевски миостромини, изградени сарколемата и, очевидно, "Z дискове", свързващи тънки актинови нишки към сарколемата. Възможно е миостромините да се съдържат заедно с актина в тънки нишки на "J дискове". АТФ е пряк източник на енергия не само за различни физиологични функции (мускулни контракции, нервна дейност, предаване на нервно възбуждане, процеси на секреция и др.), Но също така и за пластични процеси, протичащи в тялото (изграждане и обновяване на тъканни протеини, биологични синтези). Съществува постоянна конкуренция между тези две страни на живота - енергийното снабдяване на физиологичните функции и енергийното снабдяване на пластичните процеси. Увеличаването на специфичната функционална активност винаги е придружено от увеличаване на потреблението на АТФ и следователно намаляване на възможността за използването му за биологичен синтез. Както е известно, в тъканите на тялото, включително в мускулите, техните протеини непрекъснато се обновяват, но процесите на разграждане и синтез са строго балансирани и нивото на протеиново съдържание остава постоянно. По време на мускулната активност обновяването на протеините се инхибира и колкото повече, толкова повече намалява съдържанието на АТФ в мускулите. Следователно, по време на упражнения с максимална и субмаксимална интензивност, когато ресинтезът на АТФ се извършва предимно анаеробно и най-малко пълно, обновяването на протеините ще бъде инхибирано по-значително, отколкото по време на работа със средна и умерена интензивност, когато преобладават енергийно високоефективните процеси на респираторно фосфорилиране. Инхибирането на обновяването на протеините е следствие от липсата на АТФ, който е необходим както за процеса на разграждане, така и (по-специално) за процеса на техния синтез. Поради това при интензивна мускулна дейност се нарушава балансът между разграждането и синтеза на протеините, като първите преобладават над вторите. Съдържанието на протеини в мускулите леко намалява, а съдържанието на полипептиди и азотсъдържащи вещества от непротеинова природа се увеличава. Някои от тези вещества, както и някои нискомолекулни протеини, напускат мускулите в кръвта, където съответно се увеличава съдържанието на протеинов и небелтъчен азот. В този случай протеинът може да се появи и в урината. Всички тези промени са особено значими по време на силови упражнения с висока интензивност. При интензивна мускулна активност образуването на амоняк също се увеличава в резултат на дезаминиране на част от аденозинмонофосфорната киселина, която няма време да се ресинтезира в АТФ, както и поради разцепването на амоняка от глутамин, което се засилва при влиянието на повишеното съдържание на неорганични фосфати в мускулите, активиращи ензима глутаминаза. Увеличава се съдържанието на амоняк в мускулите и кръвта. Елиминирането на получения амоняк може да се осъществи главно по два начина: свързването на амоняка с глутаминовата киселина до образуване на глутамин или образуването на урея. И двата процеса обаче изискват участието на АТФ и следователно (поради намаляване на съдържанието му) изпитват затруднения по време на интензивна мускулна дейност. По време на мускулна активност със среден и умерен интензитет, когато ресинтезът на АТФ се извършва поради респираторно фосфорилиране, елиминирането на амоняка е значително повишено. Съдържанието му в кръвта и тъканите намалява, а образуването на глутамин и урея се увеличава. Поради липсата на АТФ по време на мускулна активност с максимална и субмаксимална интензивност, редица други биологични синтези също са затруднени. По-специално, синтезът на ацетилхолин в двигателните нервни окончания, което влияе отрицателно върху предаването на нервно възбуждане към мускулите.
2. Биохимични промени в тялото на бойните изкуства
Енергийните нужди на тялото (работещите мускули) се задоволяват, както е известно, по два основни начина - анаеробни и аеробни. Съотношението на тези два пътя на производство на енергия варира при различните упражнения. При извършване на всяко упражнение практически работят и трите енергийни системи: анаеробните фосфагенни (алактатни) и млечнокиселите (гликолитични) и аеробните (кислородни, окислителни) „зони” на тяхното действие частично се припокриват. Следователно е трудно да се изолира „нетният" принос на всяка от енергийните системи, особено когато работят за относително кратка максимална продължителност. В това отношение „съседните" системи по отношение на енергийната мощност (област на действие) са често се комбинират по двойки, фосфаген с лактацид, лактацид с кислород. Системата, чийто енергиен принос е по-голям, е посочена първа. Според относителното натоварване на анаеробната и аеробната енергийна система всички упражнения могат да бъдат разделени на анаеробни и аеробни. Първият - с преобладаване на анаеробния, вторият - аеробния компонент на производството на енергия.Водещото качество при изпълнение на анаеробни упражнения е силата (скоростно-силови възможности), при изпълнение на аеробни упражнения - издръжливост. Съотношението на различните системи за производство на енергия до голяма степен определя характера и степента на промени в дейността на различните физиологични системи, които осигуряват изпълнението на различни упражнения.
Има три групи анаеробни упражнения: - максимална анаеробна мощност (анаеробна мощност); - близо до максимална анаеробна мощност; - субмаксимална анаеробна мощност (анаеробно-аеробна мощност). Упражненията с максимална анаеробна мощност (анаеробна мощност) са упражнения с почти изключително анаеробен метод за доставяне на енергия на работещите мускули: анаеробният компонент в общото производство на енергия варира от 90 до 100%. Осигурява се главно от фосфагенната енергийна система (ATP + CP) с известно участие на системата на млечната киселина (гликолитичната). Рекордната максимална анаеробна мощност, развита от изключителни спортисти по време на спринт, достига 120 kcal/min. Възможната максимална продължителност на такива упражнения е няколко секунди. Укрепването на активността на вегетативните системи става постепенно по време на работа. Поради кратката продължителност на анаеробните упражнения, по време на тяхното изпълнение функциите на кръвообращението и дишането нямат време да достигнат възможния си максимум. По време на максимално анаеробно упражнение спортистът или не диша изобщо, или успява да завърши само няколко дихателни цикъла. Съответно, "средната" белодробна вентилация не надвишава 20-30% от максималната. Сърдечната честота се увеличава още преди началото (до 140-150 удара/мин) и продължава да расте по време на упражнението, като достига най-високата си стойност веднага след финала - 80-90% от максимума (160-180 удара/мин).
Тъй като енергийната основа на тези упражнения са анаеробни процеси, засилването на дейността на кардио-респираторната (пренос на кислород) система практически няма значение за енергийното осигуряване на самото упражнение. Концентрацията на лактат в кръвта по време на работа се променя много малко, въпреки че в работещите мускули може да достигне 10 mmol/kg или дори повече в края на работата. Концентрацията на лактат в кръвта продължава да се повишава няколко минути след спиране на работата и достига максимум 5-8 mmol/l. Преди извършване на анаеробни упражнения концентрацията на глюкоза в кръвта леко се повишава. Преди и в резултат на прилагането им, концентрацията на катехоламини (адреналин и норепинефрин) и растежен хормон в кръвта се увеличава много значително, но концентрацията на инсулин леко намалява; концентрациите на глюкагон и кортизол не се променят забележимо. Водещите физиологични системи и механизми, които определят спортните резултати при тези упражнения, са централната нервна регулация на мускулната дейност (координация на движенията с проява на голяма мускулна сила), функционалните свойства на нервно-мускулната система (скоростно-силови), капацитетът и мощността на фосфагенната енергийна система на работещите мускули.
Упражнения с близка до максималната анаеробна мощност (смесена анаеробна мощност) са упражнения с преобладаващо анаеробно енергоснабдяване на работещите мускули. Анаеробният компонент в общото производство на енергия е 75-85% - отчасти се дължи на фосфагена и в по-голяма степен на млечнокиселите (гликолитични) енергийни системи. Възможната максимална продължителност на такива упражнения за изключителни спортисти варира от 20 до 50 секунди. За да се осигури енергия за тези упражнения, значително увеличаване на активността на системата за транспортиране на кислород вече играе определена енергийна роля и колкото по-голяма е, колкото по-дълго е упражнението.
По време на упражнението белодробната вентилация бързо се увеличава, така че до края на упражнението, което продължава около 1 минута, може да достигне 50-60% от максималната работна вентилация за даден спортист (60-80 l/min). Концентрацията на лактат в кръвта след физическо натоварване е много висока – до 15 mmol/l при квалифицирани спортисти. Натрупването на лактат в кръвта е свързано с много висока скорост на образуването му в работещите мускули (в резултат на интензивна анаеробна гликолиза). Концентрацията на глюкоза в кръвта е леко повишена в сравнение с условията на покой (до 100-120 mg%). Хормоналните промени в кръвта са подобни на тези, които се случват по време на упражнения с максимална анаеробна мощност.
Водещите физиологични системи и механизми, които определят спортните постижения при упражнения близо до максималната анаеробна мощност, са същите като при упражненията от предишната група и в допълнение мощността на млечнокиселата (гликолитична) енергийна система на работещите мускули. Упражненията с субмаксимална анаеробна мощност (анаеробно-аеробна мощност) са упражнения с преобладаване на анаеробния компонент на енергоснабдяването на работещите мускули. В общото енергопроизводство на организма тя достига 60-70% и се осигурява основно от млечнокисела (гликолитична) енергийна система. Значителен дял от енергийното снабдяване за тези упражнения принадлежи на кислородната (окислителна, аеробна) енергийна система. Възможната максимална продължителност на състезателните упражнения за изявени спортисти е от 1 до 2 минути. Силата и максималната продължителност на тези упражнения са такива, че в процеса на тяхното изпълнение показателите за ефективност. Системата за транспортиране на кислород (сърдечен ритъм, сърдечен дебит, PV, скорост на потребление на O2) може да бъде близо до или дори да достигне максималните стойности за даден спортист. Колкото по-дълго е упражнението, толкова по-високи са тези показатели на финалната линия и толкова по-голям е делът на производството на аеробна енергия по време на упражнението. След тези упражнения се регистрира много висока концентрация на лактат в работещите мускули и кръв - до 20-25 mmol/l. По този начин тренировъчната и състезателната дейност на спортистите по бойни изкуства се провежда при приблизително максимално натоварване на мускулите на спортистите. В същото време енергийните процеси, протичащи в тялото, се характеризират с факта, че поради кратката продължителност на анаеробните упражнения по време на тяхното изпълнение функциите на кръвообращението и дишането нямат време да достигнат възможния максимум. По време на максимално анаеробно упражнение спортистът или не диша изобщо, или успява да завърши само няколко дихателни цикъла. Съответно, "средната" белодробна вентилация не надвишава 20-30% от максималната.
Човек извършва физически упражнения и изразходва енергия, използвайки нервно-мускулната система. Нервно-мускулната система е набор от двигателни единици. Всяка двигателна единица включва двигателен неврон, аксон и набор от мускулни влакна. Количеството MU остава непроменено при хората. Количеството MV в мускула е възможно и може да се променя по време на тренировка, но не повече от 5%. Следователно този фактор за растежа на мускулната функционалност няма практическо значение. Вътре в CF възниква хиперплазия (увеличаване на броя на елементите) на много органели: миофибрили, митохондрии, саркоплазмен ретикулум (SRR), гликогенови глобули, миоглобин, рибозоми, ДНК и др. Броят на капилярите, обслужващи CF, също се променя. Миофибрилата е специализиран органел на мускулното влакно (клетка). Има приблизително еднакво напречно сечение при всички животни. Състои се от последователно свързани саркомери, всеки от които включва актинови и миозинови нишки. Между актиновите и миозиновите нишки могат да се образуват мостове и с изразходването на енергията, съдържаща се в АТФ, мостовете могат да се въртят, т.е. контракция на миофибрилите, контракция на мускулните влакна, мускулна контракция. Мостовете се образуват в присъствието на калциеви йони и АТФ молекули в саркоплазмата. Увеличаването на броя на миофибрилите в мускулното влакно води до увеличаване на неговата сила, скорост на свиване и размер. Заедно с растежа на миофибрилите растат и други органели, обслужващи миофибрилите, например саркоплазмения ретикулум. Саркоплазменият ретикулум е мрежа от вътрешни мембрани, която образува везикули, тубули и цистерни. В MV SPR образува цистерни; калциевите йони (Ca) се натрупват в тези цистерни. Предполага се, че гликолитичните ензими са прикрепени към SPR мембраните, поради което при спиране на достъпа на кислород се получава значително подуване на каналите. Това явление се свързва с натрупването на водородни йони (Н), които причиняват частично разрушаване (денатуриране) на протеиновите структури и добавяне на вода към радикалите на протеиновите молекули. За механизма на мускулна контракция скоростта на изпомпване на Ca от саркоплазмата е от основно значение, тъй като това осигурява процеса на мускулна релаксация. Натриевата, калиевата и калциевата помпи са вградени в SPR мембраните, така че може да се предположи, че увеличаването на повърхността на SPR мембраните по отношение на масата на миофибрилите трябва да доведе до увеличаване на скоростта на MV релаксация.
Следователно, увеличаването на максималната скорост или скорост на мускулна релаксация (интервалът от време от края на електрическото активиране на мускула до момента, в който механичното напрежение в него спадне до нула) трябва да показва относително увеличение на мембраните на SPR. Поддържането на максимално темпо се осигурява от резервите в MV на ATP, KrF, масата на миофибриларните митохондрии, масата на саркоплазмените митохондрии, масата на гликолитичните ензими и буферния капацитет на съдържанието на мускулните влакна и кръвта.
Всички тези фактори влияят върху процеса на енергийно снабдяване на мускулната контракция, но способността за поддържане на максимално темпо трябва да зависи предимно от митохондриите на SPR. Чрез увеличаване на количеството на окислителната MV или, с други думи, аеробния капацитет на мускула, продължителността на упражнението при максимална мощност се увеличава. Това се дължи на факта, че поддържането на концентрацията на CrF по време на гликолиза води до подкисляване на MV, инхибиране на процесите на потребление на АТФ поради конкуренцията на Н йони с Са йони в активните центрове на миозиновите глави. Следователно процесът на поддържане на концентрацията на CrF, с преобладаване на аеробните процеси в мускулите, става все по-ефективен с изпълнението на упражнението. Важно е също така, че митохондриите активно абсорбират водородни йони, поради което при извършване на краткотрайни екстремни упражнения (10-30 s) тяхната роля е по-ограничена до буфериране на подкиселяването на клетките. По този начин адаптирането към мускулната работа се осъществява чрез работата на всяка клетка на спортиста, въз основа на енергийния метаболизъм по време на живота на клетката. Основата на този процес е консумацията на АТФ по време на взаимодействието на водородни и калциеви йони.
Увеличаването на развлекателната стойност на битките включва значително увеличаване на активността на битката с едновременно увеличаване на броя на изпълняваните технически действия. Като се има предвид това, възниква реален проблем, свързан с факта, че при повишена интензивност на състезателен мач на фона на прогресираща физическа умора ще настъпи временна автоматизация на двигателните умения на спортиста.
В спортната практика това обикновено се проявява през второто полувреме на състезателен мач, проведен с висока интензивност. В този случай (особено ако спортистът няма много високо ниво на специална издръжливост) се наблюдават значителни промени в рН на кръвта (под 7,0 конвенционални единици), което показва изключително неблагоприятна реакция на спортиста към работа с такава интензивност. Известно е, че например стабилно нарушаване на ритмичната структура на двигателните умения на борец при извършване на хвърляне на заден огън започва с нивото на физическа умора при стойности на рН на кръвта под 7,2 арб. единици
В тази връзка има два възможни начина за повишаване на стабилността на двигателните умения на бойните изкуства: а) повишаване на нивото на специална издръжливост до такава степен, че да могат да провеждат битка с всякаква интензивност без изразена физическа умора (реакция натоварването не трябва да води до ацидотични промени под стойностите на pH, равни на 7,2 конвенционални единици); б) осигурява стабилно проявление на двигателните умения във всякакви екстремни ситуации на екстремна физическа активност при стойности на рН на кръвта, достигащи 6,9 конвенционални стойности. единици В рамките на първото направление бяха проведени доста голям брой специални изследвания, които определиха реалните начини и перспективи за решаване на проблема с ускореното обучение на специална издръжливост при спортисти по бойни изкуства. По втория проблем към днешна дата няма реални, практически значими разработки.
4. Проблемът с възстановяването в спорта
Едно от най-важните условия за интензифициране на тренировъчния процес и по-нататъшно повишаване на спортните постижения е широкото и систематично използване на възстановителни средства. Рационалното възстановяване е от особено значение при екстремни и близки до максимални физически и психически натоварвания – задължителни спътници на тренировки и състезания в съвременния спорт. Очевидно използването на система от възстановителни средства налага ясно да се класифицират процесите на възстановяване в условията на спортна дейност.
Спецификата на възстановителните промени, обусловена от естеството на спортната дейност, обема и интензивността на тренировъчните и състезателни натоварвания и общия режим, определя конкретни мерки, насочени към възстановяване на ефективността. Н. И. Волков идентифицира следните видове възстановяване при спортисти: текущо (наблюдение по време на работа), спешно (след края на натоварването) и забавено (много часове след завършване на работата), както и след хронично пренапрежение (т.нар. възстановяване от стрес). Трябва да се отбележи, че изброените реакции се извършват на фона на периодично възстановяване поради потребление на енергия при нормални условия на живот.
Неговият характер до голяма степен се определя от функционалното състояние на тялото. Ясното разбиране на динамиката на възстановителните процеси в условията на спортна дейност е необходимо за организиране на рационалното използване на възстановителните средства. По този начин функционалните промени, които се развиват в процеса на продължаващо възстановяване, са насочени към осигуряване на повишени енергийни нужди на тялото, за компенсиране на увеличеното потребление на биологична енергия в процеса на мускулна дейност. Метаболитните трансформации заемат централно място във възстановяването на енергийните разходи.
Съотношението на разхода на енергия на тялото и възстановяването му по време на работа позволява да се раздели физическата активност на 3 диапазона: 1) натоварвания, при които е достатъчна аеробна подкрепа за работа; 2) натоварвания, при които наред с аеробната поддръжка на работата се използват анаеробни източници на енергия, но границата на увеличаване на доставката на кислород към работещите мускули все още не е надвишена; 3) натоварвания, при които енергийните нужди надвишават възможностите за текущо възстановяване, което е придружено от бързо развиваща се умора. В някои спортове, за да се оцени ефективността на рехабилитационните мерки, е препоръчително да се анализират различни показатели на нервно-мускулната система и да се използват психологически тестове. Използването в практиката на работа със спортисти от висок клас на задълбочени прегледи с помощта на обширен набор от инструменти и методи ни позволява да оценим ефективността на предишни рехабилитационни мерки и да определим тактиката на следващите. Тестовете за възстановяване изискват поетапни прегледи, извършвани в седмични или месечни цикли на обучение. Честотата на тези прегледи и методите на изследване се определят от лекаря и треньора в зависимост от вида на спорта, естеството на натоварванията в даден тренировъчен период, използваните възстановителни средства и индивидуалните характеристики на спортиста.
5 . Характеристики на метаболитните състояния при хора по време на мускулна дейност
Състоянието на метаболизма в човешкото тяло се характеризира с голям брой променливи. В условия на интензивна мускулна дейност най-важният фактор, от който зависи метаболитното състояние на организма, е приложението в областта на енергийния метаболизъм. За количествено определяне на метаболитните състояния при хора по време на мускулна работа се предлага да се използват три типа критерии: а) критерии за мощност, отразяващи скоростта на преобразуване на енергията в аеробни и анаеробни процеси; б) критерии за капацитет, характеризиращи енергийните резерви на тялото или общия обем на метаболитните промени, настъпили по време на работа; в) критерии за ефективност, които определят степента, в която енергията на аеробните и анаеробните процеси се използва при извършване на мускулна работа. Промените в силата и продължителността на упражненията имат различен ефект върху аеробния и анаеробния метаболизъм. Такива показатели за мощността и капацитета на аеробния процес, като размера на белодробната вентилация, нивото на консумация на кислород и прием на кислород по време на работа, систематично се увеличават с продължителността на упражнението при всяка избрана стойност на мощността. Тези показатели нарастват значително с увеличаване на интензивността на работа във всички интервали от време на упражнението. Индикаторите за максимално натрупване на млечна киселина в кръвта и общия кислороден дълг, които характеризират капацитета на анаеробните енергийни източници, се променят малко при изпълнение на упражнения с умерена мощност, но се увеличават значително с увеличаване на продължителността на работа при по-интензивни упражнения.
Интересно е да се отбележи, че при най-ниска мощност на натоварване, където съдържанието на млечна киселина в кръвта остава на постоянно ниво от около 50-60 mg, е практически невъзможно да се открие лактатната фракция на кислородния дълг; Няма прекомерно отделяне на въглероден диоксид, свързано с разрушаването на кръвните бикарбонати по време на натрупването на млечна киселина. Може да се предположи, че отбелязаното ниво на натрупване на млечна киселина в кръвта все още не надвишава тези прагови стойности, над които се наблюдава стимулиране на окислителните процеси, свързани с елиминирането на лактатния кислороден дълг. Индикаторите на аеробния метаболизъм след кратък период на забавяне (около 1 минута), свързан с тренировка, показват системно увеличение с увеличаване на времето за упражнение.
По време на периода на разработка се наблюдава изразено засилване на анаеробните реакции, водещи до образуване на млечна киселина. Увеличаването на силата на упражненията е придружено от пропорционално увеличаване на аеробните процеси. Увеличаване на интензивността на аеробните процеси с увеличаване на мощността се установява само при упражнения с продължителност над 0,5 минути. При извършване на интензивни краткотрайни упражнения се наблюдава намаляване на аеробния метаболизъм. Увеличаване на размера на общия кислороден дълг поради образуването на лактатна фракция и появата на излишно отделяне на въглероден диоксид се открива само при тези упражнения, чиято мощност и продължителност са достатъчни за натрупване на млечна киселина над 50-60 mg %. При изпълнение на упражнения с ниска мощност промените в показателите на аеробните и анаеробните процеси показват обратна посока; с увеличаване на мощността промените в тези процеси се променят към еднопосочни.
В динамиката на показателите за скоростта на консумация на кислород и „излишното“ отделяне на въглероден диоксид по време на тренировка се открива фазово изместване; по време на периода на възстановяване след края на работата се случва синхронизиране на промените в тези показатели. Промените в консумацията на кислород и нивата на млечна киселина в кръвта с увеличаване на времето за възстановяване след интензивни упражнения ясно показват фазови разлики. Проблемът с умората в биохимията на спорта е един от най-трудните и все още далеч от своето решение. В най-общия си вид умората може да се определи като състояние на тялото, което възниква в резултат на продължителна или усилена дейност и се характеризира с намаляване на работоспособността. Субективно се възприема от човек като чувство на локална умора или обща умора. Дългосрочните проучвания позволяват да се разделят биохимичните фактори, които ограничават ефективността, на три групи, свързани помежду си.
Това са, на първо място, биохимични промени в централната нервна система, причинени както от самия процес на двигателно възбуждане, така и от проприоцептивни импулси от периферията. На второ място, това са биохимични промени в скелетните мускули и миокарда, причинени от тяхната работа и трофични промени в нервната система. Трето, това са биохимични промени във вътрешната среда на тялото, зависещи както от процесите, протичащи в мускулите, така и от влиянието на нервната система. Общи характеристики на умората са дисбаланс на фосфатни макроерги в мускулите и мозъка, както и намаляване на АТФазната активност и коефициента на фосфорилиране в мускулите. Но умората, свързана с работа с висока интензивност и продължителна работа, има и някои специфични особености. В допълнение, биохимичните промени по време на умора, причинена от краткотрайна мускулна активност, се характеризират със значително по-голям градиент, отколкото по време на мускулна активност с умерена интензивност, но продължителността е близо до границата. Трябва да се подчертае, че рязкото намаляване на въглехидратните резерви на тялото, макар и от голямо значение, не играе решаваща роля за ограничаване на работоспособността. Най-важният фактор, ограничаващ производителността, е нивото на АТФ както в самите мускули, така и в централната нервна система.
В същото време не можете да пренебрегнете биохимичните промени в други органи, по-специално в миокарда. При интензивна краткотрайна работа нивото на гликоген и креатин фосфат в него не се променя, но се повишава активността на окислителните ензими. При продължителна работа може да има намаляване както на нивото на гликоген и креатин фосфат, така и на ензимната активност. Това е придружено от промени в ЕКГ, показващи дистрофични процеси, най-често в лявата камера и по-рядко в предсърдията. По този начин умората се характеризира с дълбоки биохимични промени както в централната нервна система, така и в периферията, предимно в мускулите. Освен това степента на биохимичните промени в последния може да се промени с повишена производителност, причинена от ефекта върху централната нервна система. I.M. пише за централната нервна природа на умората още през 1903 г. Сеченов. Оттогава данните за ролята на централното инхибиране в механизма на умората нарастват. Наличието на дифузно инхибиране по време на умора, причинена от продължителна мускулна активност, не може да бъде съмнено. Той се развива в централната нервна система и се развива в нея чрез взаимодействието на центъра и периферията с водеща роля на първата. Умората е следствие от промени в организма, причинени от интензивна или продължителна дейност, и защитна реакция, която предотвратява преминаването през линията на функционални и биохимични нарушения, които са опасни за организма и застрашават неговото съществуване.
Определена роля в механизма на умората играят и нарушенията в метаболизма на протеините и нуклеиновите киселини в нервната система. При продължително бягане или плуване с товар, което причинява значителна умора, се наблюдава намаляване на нивата на РНК в двигателните неврони, докато при продължителна, но неуморителна работа то не се променя или се повишава. Тъй като химията и по-специално активността на мускулните ензими се регулират от трофичните влияния на нервната система, може да се предположи, че промените в химичния статус на нервните клетки по време на развитието на защитно инхибиране, причинено от умора, водят до промени в трофичния центробежен импулси, водещи до нарушения в регулацията на мускулната химия.
Тези трофични влияния очевидно се осъществяват чрез движението на биологично активни вещества по аксоплазмата на еферентните влакна, описани от P. Weiss. По-специално, от периферните нерви е изолирано протеиново вещество, което е специфичен инхибитор на хексокиназата, подобен на инхибитора на този ензим, секретиран от предната хипофизна жлеза. По този начин умората се развива чрез взаимодействие на централни и периферни механизми с водещо и интегриращо значение на първите. Свързва се както с промени в нервните клетки, така и с рефлексни и хуморални влияния от периферията. Биохимичните промени при умора могат да бъдат генерализирани, придружени от общи промени във вътрешната среда на тялото и нарушения в регулацията и координацията на различни физиологични функции (при продължителна физическа активност, включваща значителна мускулна маса). Тези промени могат да бъдат и от по-локален характер, да не са придружени от значителни общи промени, но да се ограничават само до работещите мускули и съответните групи нервни клетки и центрове (по време на краткотрайна работа с максимална интензивност или дългосрочна работа на ограничен брой мускули).
Умората (и особено чувството на умора) е защитна реакция, която предпазва организма от прекомерни степени на функционално изтощение, които са животозастрашаващи. Едновременно с това тренира физиологични и биохимични компенсаторни механизми, създавайки предпоставки за възстановителни процеси и допълнително повишавайки функционалността и работоспособността на организма. По време на почивка след мускулна работа нормалните съотношения на биологични съединения се възстановяват както в мускулите, така и в тялото като цяло. Ако по време на мускулна работа доминират катаболните процеси, необходими за снабдяване с енергия, тогава по време на почивка преобладават анаболните процеси. Анаболните процеси изискват разход на енергия под формата на АТФ, поради което най-изразените промени се откриват в областта на енергийния метаболизъм, тъй като през периода на почивка АТФ непрекъснато се изразходва и следователно резервите на АТФ трябва да бъдат възстановени. Анаболните процеси по време на почивка се дължат на катаболни процеси, настъпили по време на работа. По време на почивка АТФ, креатин фосфат, гликоген, фосфолипиди и мускулни протеини се ресинтезират, водно-електролитният баланс на тялото се нормализира и увредените клетъчни структури се възстановяват. В зависимост от общата посока на биохимичните промени в организма и времето, необходимо за процесите на разделяне, се разграничават два вида възстановителни процеси - спешно и изоставено възстановяване. Спешното възстановяване продължава от 30 до 90 минути след работа. По време на периода на спешно възстановяване се елиминират продуктите от анаеробно разлагане, натрупани по време на работа, предимно млечна киселина и кислороден дълг. След приключване на работа, консумацията на кислород продължава да бъде повишена в сравнение със състоянието на покой. Тази излишна консумация на кислород се нарича кислороден дълг. Кислородният дълг винаги е по-голям от кислородния дефицит и колкото по-висока е интензивността и продължителността на работа, толкова по-значима е тази разлика.
По време на почивка консумацията на АТФ за мускулни контракции спира и съдържанието на АТФ в митохондриите се увеличава през първите секунди, което показва прехода на митохондриите в активно състояние. Концентрацията на АТФ се увеличава, повишавайки нивото преди работа. Повишава се и активността на окислителните ензими. Но активността на гликогенфосфорилазата рязко намалява. Млечната киселина, както вече знаем, е крайният продукт от разграждането на глюкозата при анаеробни условия. В началния момент на почивка, когато остава повишена консумация на кислород, се увеличава доставката на кислород към окислителните системи на мускулите. В допълнение към млечната киселина, други метаболити, натрупани по време на работа, също са обект на окисление: янтарна киселина, глюкоза; и в по-късните етапи на възстановяване, мастни киселини. Възстановяването на забавяне продължава дълго след приключване на работата. На първо място, той засяга процесите на синтез на структури, изразходвани по време на мускулната работа, както и възстановяването на йонния и хормоналния баланс в тялото. По време на възстановителния период се натрупват запаси от гликоген в мускулите и черния дроб; тези процеси на възстановяване протичат в рамките на 12-48 часа. Млечната киселина, навлизаща в кръвта, навлиза в чернодробните клетки, където първо се осъществява синтеза на глюкоза, а глюкозата е директният строителен материал за гликоген синтетазата, която катализира синтеза на гликоген. Процесът на ресинтеза на гликоген има фазов характер, който се основава на феномена на суперкомпенсация. Суперкомпенсация (свръхвъзстановяване) е превишаването на запасите от енергийни вещества през периода на почивка до работното ниво. Суперкомпенсацията е приемливо явление. Съдържанието на гликоген, което е намаляло след работа, се увеличава по време на почивка не само до първоначалното ниво, но и до по-високо ниво. След това има спад до първоначалното (до работното) ниво и дори малко по-ниско, а след това има вълнообразно връщане към първоначалното ниво.
Продължителността на фазата на суперкомпенсация зависи от продължителността на работата и дълбочината на биохимичните промени, които причинява в тялото. Мощната краткотрайна работа предизвиква бързо начало и бързо завършване на фазата на суперкомпенсация: когато интрамускулните резерви на гликоген се възстановят, фазата на суперкомпенсация се открива след 3-4 часа и завършва след 12 часа. След продължителна работа с умерена мощност, суперкомпенсацията на гликогена настъпва след 12 часа и завършва между 48 и 72 часа след края на работата. Законът за суперкомпенсацията е валиден за всички биологични съединения и структури, които в една или друга степен се изразходват или разрушават по време на мускулна дейност и се ресинтезират по време на почивка. Те включват: креатин фосфат, структурни и ензимни протеини, фосфолипиди, клетъчна оргонела (митохондрии, лизозоми). След ресинтеза на енергийните резерви на организма значително се засилват процесите на ресинтез на фосфолипиди и протеини, особено след тежка силова работа, която е придружена от значителното им разпадане. Възстановяването на нивото на структурните и ензимните протеини става в рамките на 12-72 часа. При извършване на работа, свързана със загуба на вода, резервите от вода и минерални соли трябва да се попълват през периода на възстановяване. Основният източник на минерални соли е храната.
6 . Биохимичен контрол в бойните изкуства
При интензивна мускулна дейност в мускулите се образуват големи количества млечна и пирогроздена киселина, които дифундират в кръвта и могат да причинят метаболитна ацидоза на организма, която води до мускулна умора и е съпроводена с мускулни болки, световъртеж и гадене. Такива метаболитни промени са свързани с изчерпването на буферните резерви на организма. Тъй като състоянието на буферните системи на тялото е важно за проявата на висока физическа работоспособност, показателите на CBS се използват в спортната диагностика. Показателите на CBS, които обикновено са относително постоянни, включват: - pH на кръвта (7,35-7,45); - pCO2 - парциално налягане на въглеродния диоксид (H2CO3 + CO2) в кръвта (35 - 45 mm Hg); - 5B - стандартен кръвен плазмен бикарбонат HSOd, който при пълно насищане на кръвта с кислород е 22-26 meq/l; - BB - буферни основи на цяла кръв или плазма (43 - 53 meq/l) - показател за капацитета на цялата буферна система от кръв или плазма; - L/86 - нормални буферни основи на цяла кръв при физиологични стойности на pH и CO2 на алвеоларен въздух; - BE - излишък на основа, или алкален резерв (от - 2,4 до +2,3 meq/l) - индикатор за излишък или дефицит на буфер. Индикаторите на CBS отразяват не само промените в буферните системи на кръвта, но и състоянието на дихателната и отделителната система на тялото. Състоянието на киселинно-алкалния баланс (ABC) в организма се характеризира с постоянно рН на кръвта (7,34-7,36).
Установена е обратна корелация между динамиката на съдържанието на лактат в кръвта и промените в рН на кръвта. Чрез промяна на показателите на ABS по време на мускулна активност е възможно да се наблюдава реакцията на тялото към физическа активност и растежа на годността на спортиста, тъй като с биохимичния контрол на ABS може да се определи един от тези показатели. Активната реакция на урината (pH) е пряко зависима от киселинно-алкалното състояние на тялото. При метаболитна ацидоза киселинността на урината се повишава до pH 5, а при метаболитна алкалоза намалява до pH 7. Таблица. Фигура 3 показва посоката на промените в стойностите на pH на урината по отношение на показателите за киселинно-алкалното състояние на плазмата. По този начин борбата като спорт се характеризира с висока интензивност на мускулната активност. В тази връзка е важно да се контролира обмяната на киселини в тялото на спортиста. Най-информативният показател за ACS е стойността на BE - алкален резерв, който нараства с повишаване на квалификацията на спортистите, особено на тези, които се специализират в скоростно-силовите спортове.
Заключение
В заключение можем да кажем, че тренировъчната и състезателната дейност на бойните изкуства протича при приблизително максимално натоварване на мускулите на спортистите. В същото време енергийните процеси, протичащи в тялото, се характеризират с факта, че поради кратката продължителност на анаеробните упражнения по време на тяхното изпълнение функциите на кръвообращението и дишането нямат време да достигнат възможния максимум. По време на максимално анаеробно упражнение спортистът или не диша изобщо, или успява да завърши само няколко дихателни цикъла. Съответно, "средната" белодробна вентилация не надвишава 20-30% от максималната. Умората при състезателни и тренировъчни дейности на спортистите по бойни изкуства възниква поради почти максимално натоварване на мускулите през целия период на битката.
В резултат на това нивото на pH в кръвта се повишава, реакцията на спортиста и неговата устойчивост на атаки от врага се влошават. За намаляване на умората се препоръчва използването на гликолитични анаеробни натоварвания в тренировъчния процес. Процесът на следа, създаден от доминиращия фокус, може да бъде доста устойчив и инертен, което прави възможно поддържането на възбуда дори когато източникът на дразнене е премахнат.
След края на мускулната работа започва период на възстановяване или период след работа. Характеризира се със степента на промяна във функциите на тялото и времето, необходимо за възстановяването им до първоначалното ниво. Проучването на периода на възстановяване е необходимо, за да се оцени тежестта на определена работа, да се определи нейното съответствие с възможностите на тялото и да се определи продължителността на необходимата почивка. Биохимичната основа на двигателните умения на бойните изкуства е пряко свързана с проявата на силовите способности, които включват динамична, експлозивна и изометрична сила. Адаптирането към мускулната работа се осъществява чрез работата на всяка клетка на спортиста, въз основа на енергийния метаболизъм по време на живота на клетката. Основата на този процес е консумацията на АТФ по време на взаимодействието на водородни и калциеви йони. Бойните изкуства, като спорт, се характеризират с висока интензивност на мускулната активност. В тази връзка е важно да се контролира обмяната на киселини в тялото на спортиста. Най-информативният показател за ACS е стойността на BE - алкален резерв, който нараства с повишаване на квалификацията на спортистите, особено на тези, които се специализират в скоростно-силовите спортове.
Библиография
1. Волков Н.И. Биохимия на мускулната дейност. - М.: Олимпийски спорт, 2001.
2. Волков Н.И., Олейников В.И. Биоенергия на спорта. - М: Съветски спорт, 2011.
3. Максимов Д.В., Селуянов В.Н., Табаков С.Е. Физическа подготовка на бойни изкуства. - М: TVT Division, 2011.
Публикувано на Allbest.ru
Подобни документи
Мускулно-скелетна система на цитоплазмата. Структура и химичен състав на мускулната тъкан. Функционална биохимия на мускулите. Биоенергийни процеси при мускулна дейност. Биохимия на физическите упражнения. Биохимични промени в мускулите по време на патология.
ръководство за обучение, добавено на 19.07.2009 г
Същността на концепцията и основните функции на мускулната дейност. Етап на възстановяване на човешкото тяло. Индикатори за възстановяване и средства, които ускоряват процеса. Основните физиологични характеристики на бързото пързаляне с кънки.
тест, добавен на 30.11.2008 г
Биохимичен мониторинг на тренировъчния процес. Видове лабораторен контрол. Система за енергоснабдяване на тялото. Характеристики на храненето за спортисти. Начини за преобразуване на енергия. Степен на обучение, основни видове адаптация, техните характеристики.
дисертация, добавена на 22.01.2018 г
Мускулите като органи на човешкото тяло, състоящи се от мускулна тъкан, способна да се съкращава под въздействието на нервни импулси, тяхната класификация и видове, функционална роля. Характеристики на мускулната работа на човешкото тяло, динамична и статична.
презентация, добавена на 23.04.2013 г
Скелетна мускулна маса при възрастен. Активна част от опорно-двигателния апарат. Напречно набраздени мускулни влакна. Устройството на скелетните мускули, основните групи и гладките мускули и тяхната работа. Свързани с възрастта характеристики на мускулната система.
тест, добавен на 19.02.2009 г
Биохимични анализи в клиничната медицина. Протеини на кръвната плазма. Клинична биохимия на заболявания на черния дроб, стомашно-чревния тракт, нарушения на хемостазата, анемия и кръвопреливане, захарен диабет, ендокринни заболявания.
ръководство за обучение, добавено на 19.07.2009 г
Характеристики на източниците на развитие на сърдечната мускулна тъкан, които се намират в прекордиалната мезодерма. Анализ на кардиомиоцитната диференциация. Характеристики на структурата на сърдечната мускулна тъкан. Същността на процеса на регенерация на сърдечната мускулна тъкан.
презентация, добавена на 07/11/2012
Биохимични анализи в клиничната медицина. Патохимични механизми на универсалните патологични феномени. Клинична биохимия при ревматични заболявания, заболявания на дихателната система, бъбреците и стомашно-чревния тракт. Нарушения на системата за хемостаза.
ръководство за обучение, добавено на 19.07.2009 г
Физическо и психическо развитие на детето в неонатална и кърмаческа възраст. Анатомо-физиологични особености на предучилищния период от живота. Развитие на мускулната система и скелета при деца в начална училищна възраст. Периодът на пубертета при децата.
презентация, добавена на 10/03/2015
Добре оформената и функционираща опорно-двигателна система е едно от основните условия за правилното развитие на детето. Запознаване с основните характеристики на скелетно-мускулната система при децата. Обща характеристика на гърдите на новороденото.
В спортната физиология е обичайно да се разграничава и подразделя мускулната активност на зони на мощност: максимална, субмаксимална, висока и умерена. Съществува и друго разделение на мускулната работа в зависимост от основните механизми на енергоснабдяване: в анаеробни, смесени и аеробни зони на енергоснабдяване.
Във всяка мускулна работа, на първо място, трябва да се прави разлика между нейната начална (начална) фаза и нейното продължение. Времето на началната фаза зависи от интензивността на работата: колкото по-продължителна е работата, толкова по-интензивна е началната фаза и толкова по-изразени са биохимичните промени в мускулите по време на нея.
В първите секунди на работа мускулите получават по-малко кислород от необходимото. Колкото по-висока е интензивността на работа и съответно колкото по-висока е потребността от кислород, толкова по-голям е дефицитът на кислород. Следователно, в началната фаза ресинтезата на АТФ се извършва изключително анаеробно поради реакцията на креатинкиназата и гликолизата.
Ако интензивността на мускулната работа е максимална и продължителността е съответно кратка, тогава тя завършва в тази начална фаза. В този случай нуждата от кислород няма да бъде задоволена.
Когато работите с подмаксимален интензитет, но за по-дълъг период от време, биохимичните промени в началната фаза ще бъдат по-малко драматични и самата начална фаза ще бъде съкратена. В този случай потреблението на кислород ще достигне MPC (максималните възможни стойности), но търсенето на кислород все още няма да бъде задоволено. При тези условия тялото изпитва недостиг на кислород. Значението на пътя на креатинкиназата ще намалее, гликолизата ще продължи доста интензивно, но механизмите на аеробния ресинтез на АТФ вече ще бъдат активирани. Процесът на гликолиза ще включва главно глюкоза, донесена от кръвта от черния дроб, а не глюкоза, образувана от мускулен гликоген.
При мускулна работа с още по-малък интензитет и по-голяма продължителност, след краткотрайна начална фаза, ресинтезът на АТФ преобладава по аеробния механизъм, което е следствие от установяването на истински баланс между търсенето на кислород и доставката на кислород. Има повишаване и стабилизиране на нивото на АТФ в мускулните влакна, но това ниво е по-ниско, отколкото в покой. Освен това има леко повишаване на нивата на креатин фосфат.
Ако при продължителна мускулна работа силата му рязко се увеличи, тогава се наблюдават същите явления, както в началната фаза. Увеличаването на работната мощност естествено води до увеличаване на търсенето на кислород, което не може да бъде незабавно задоволено. В резултат на това се активират анаеробни механизми за ресинтеза на АТФ.
Нека разгледаме времевата последователност на активиране на различни пътища за ресинтез на АТФ. През първите 2-3 секунди от работата на мускула, неговото енергийно снабдяване се дължи на разграждането на мускулния АТФ. От 3 до 20 секунди настъпва ресинтеза на АТФ поради разграждането на креатин фосфата. След това, 30-40 секунди от началото на мускулната работа, гликолизата достига най-голяма интензивност. Освен това процесите на окислително фосфорилиране започват да играят все по-важна роля в енергийното снабдяване (фиг. 10).
Фиг. 10. Участието на различни енергийни източници в енергийното снабдяване на мускулната активност в зависимост от нейната продължителност: 1 - разграждане на АТФ, 2 - разграждане на креатин фосфат,
3 – гликолиза, 4 – аеробно окисление
Силата на производството на аеробна енергия се оценява чрез стойността на MIC. Статистиката показва, че мъжете средно имат по-висока BMD от жените. При спортистите тази стойност е значително по-висока, отколкото при нетренирани хора. Сред спортистите от различни специалности най-високите стойности на VO2 max се наблюдават при скиори и бегачи на дълги разстояния.
Системната физическа активност води до увеличаване на броя на митохондриите в мускулните клетки и до увеличаване на броя и активността на ензимите на дихателната верига. Това създава условия за по-пълноценно използване на постъпващия кислород и по-успешно енергоснабдяване на тренираното тяло.
При редовни тренировки се увеличава броят на съдовете, доставящи кръв на мускулите. Това създава по-ефективна система за снабдяване на мускулите с кислород и глюкоза, както и премахване на отпадъчните продукти. По време на дългосрочно обучение кръвоносната и дихателната системи се адаптират по такъв начин, че кислородният дефицит, който се появява след първите упражнения, впоследствие може да бъде напълно компенсиран. Способността на мускулите да работят дълго време обикновено зависи от скоростта и ефективността на тяхното усвояване и използване на кислород.
Мускулна система и нейните функции
контракции, общ преглед на скелетните мускули)
Има два вида мускули: гладка(неволно) и набразден(произволно). Гладките мускули са разположени в стените на кръвоносните съдове и някои вътрешни органи. Те свиват или разширяват кръвоносните съдове, придвижват храната по стомашно-чревния тракт и свиват стените на пикочния мехур. Напречнонабраздените мускули са всички скелетни мускули, които осигуряват разнообразни движения на тялото. Към набраздената мускулатура се отнася и сърдечният мускул, който автоматично осигурява ритмичното функциониране на сърцето през целия живот. Основата на мускулите са протеини, съставляващи 80–85% от мускулната тъкан (с изключение на водата). Основното свойство на мускулната тъкан е контрактилност,осигурява се от контрактилни мускулни протеини - актин и миозин.
Мускулната тъкан е много сложна. Мускулът има влакнеста структура, всяко влакно е мускул в миниатюра, комбинацията от тези влакна образува мускула като цяло. мускулни влакна,от своя страна се състои от миофибрилиВсяка миофибрила е разделена на редуващи се светли и тъмни области. Тъмни области - протофибрилите се състоят от дълги вериги от молекули миозин,леките се образуват от по-тънки белтъчни нишки актина.Когато мускулът е в неконтрахирано (отпуснато) състояние, актиновите и миозиновите нишки са само частично напреднали един спрямо друг, като всяка миозинова нишка е срещуположна и заобиколена от няколко актинови нишки. По-дълбокото напредване един спрямо друг причинява скъсяване (свиване) на миофибрилите на отделните мускулни влакна и на целия мускул като цяло (фиг. 2.3).
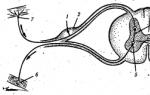
Множество нервни влакна се приближават и отклоняват от мускула (принцип на рефлексната дъга) (фиг. 2.4). Моторните (еферентни) нервни влакна предават импулси от мозъка и гръбначния мозък, привеждайки мускулите в работно състояние; сензорните влакна предават импулси в обратна посока, информирайки централната нервна система за мускулната активност. Чрез симпатиковите нервни влакна се регулират метаболитните процеси в мускулите, при което тяхната дейност се адаптира към променящите се условия на работа и различни мускулни натоварвания. Всеки мускул е пронизан от широка мрежа от капиляри, през които навлизат необходимите за функционирането на мускулите вещества и се елиминират метаболитните продукти.
Скелетни мускули.Скелетните мускули са част от структурата на опорно-двигателния апарат, прикрепени са към костите на скелета и при свиване движат отделни части на скелета и лостове. Те участват в поддържането на положението на тялото и неговите части в пространството, осигуряват движения при ходене, бягане, дъвчене, преглъщане, дишане и др., като същевременно генерират топлина. Скелетните мускули имат способността да се възбуждат под въздействието на нервни импулси. Възбуждането се извършва до контрактилни структури (миофибрили), които, свивайки се, извършват определен двигателен акт - движение или напрежение.
Ориз. 2.3. Схематично представяне на мускула.
Мускул (L) се състои от мускулни влакна (B),всяка от тях е изградена от миофибрили (IN).Миофибрила (G)съставен от дебели и тънки миофиламенти (D). Фигурата показва един саркомер, ограничен от двете страни с линии: 1 - изотропен диск, 2 - анизотропен диск, 3 - зона с по-малка анизотропия. Напречна среда на мултифибрил (4), давайки представа за шестоъгълното разпределение на дебели и тънки мултифиламенти

Ориз. 2.4. Диаграма на най-простата рефлексна дъга:
1 - аферентен (чувствителен) неврон, 2 - гръбначен възел, 3 - интерневрон, 4 .- сиво вещество на гръбначния мозък, 5 - еферентен (двигателен) неврон, 6 - моторни нервни окончания в мускулите; 7 - сетивен нерв, завършващ в кожата
Спомнете си, че всички скелетни мускули се състоят от набраздени мускули. При хората има около 600 от тях и повечето от тях са чифтни. Теглото им е 35-40% от общото телесно тегло на възрастен човек. Скелетните мускули са покрити отвън с плътна съединителнотъканна мембрана. Всеки мускул има активна част (мускулно тяло) и пасивна част (сухожилие). Мускулите се делят на дълъг, късИ широк.
Наричат се мускули, чието действие е насочено в обратна посока антагонистиеднопосочен - синергисти.Едни и същи мускули в различни ситуации могат да действат в едно и друго качество. При хората по-често се срещат вретеновидни и лентовидни. Веретенообразни мускулиразположени и функциониращи в областта на дългите костни образувания на крайниците, те могат да имат два корема (дигастрални мускули) и няколко глави (бицепс, трицепс, четириглав мускул). Лентовидни мускулиимат различна ширина и обикновено участват в корсетообразуването на стените на тялото. Мускулите с перна структура, имащи голям физиологичен диаметър поради голям брой къси мускулни структури, са много по-силни от тези мускули, в които влакната имат линейно (надлъжно) разположение. Първите се наричат силни мускули, които извършват движения с малка амплитуда, вторите се наричат сръчни мускули, които участват в движения с голяма амплитуда. Според функционалното предназначение и посоката на движение в ставите се разграничават мускулите флексориИ екстензори, адукториИ abducens, сфинктери(компресивна) и разширители.
Мускулна силаопределя се от теглото на товара, който той може да повдигне на определена височина (или може да задържи при максимално възбуждане), без да променя дължината си. Силата на мускула зависи от сумата на силите на мускулните влакна и тяхната съкратимост; върху броя на мускулните влакна в мускула и броя на функционалните единици,едновременно възбудени, когато се развие напрежение; от начална мускулна дължина(предварително разтегнатият мускул развива по-голяма сила); от условия на взаимодействие с костите на скелета.
Контрактилитетмускул се характеризира със своята абсолютна сила,тези. сила на 1 cm 2 напречно сечение на мускулни влакна. За да се изчисли този показател, мускулната сила се разделя на площ неговия физиологичен диаметър(т.е. сумата от площите на всички мускулни влакна, които изграждат мускула). Например: средният човек има силата (на 1 cm 2 мускулно напречно сечение) на коремния мускул. - 6,24; екстензори на врата - 9,0; трицепс брахии мускул - 16,8 кг.
Централната нервна система регулира силата на мускулната контракция, като променя броя на функционалните единици, участващи едновременно в контракция, както и честотата на импулсите, изпращани към тях. Увеличаването на честотата на импулса води до повишаване на напрежението.
Мускулна работа.По време на процеса на мускулно съкращение потенциалната химическа енергия се преобразува в потенциална механична енергия на напрежение и кинетична енергия на движение. Има разграничение между вътрешна и външна работа. Вътрешната работа е свързана с триене в мускулното влакно по време на неговото съкращение. Външната работа се проявява при преместване на собственото тяло, товар или отделни части на тялото (динамична работа) в пространството. Характеризира се с коефициента на ефективност (ефективност) на мускулната система, т.е. съотношението на извършената работа към общия разход на енергия (за човешките мускули ефективността е 15-20%, за физически развитите, обучени хора тази цифра е малко по-висока).
При статични усилия (без движение) можем да говорим не за работа като такава от гледна точка на физиката, а за работа, която трябва да се оцени чрез физиологичните енергийни разходи на тялото.
Мускулите като орган.Като цяло мускулът като орган е сложно структурно образувание, което изпълнява определени функции и се състои от 72-80% вода и 16-20% плътна материя. Мускулните влакна се състоят от миофибрили с клетъчни ядра, рибозоми, митохондрии, саркоплазмен ретикулум, чувствителни нервни образувания - проприорецептори и други функционални елементи, които осигуряват протеинов синтез, окислително фосфорилиране и ресинтез на аденозинтрифосфорна киселина, транспорт на вещества в мускулната клетка и др. по време на функционирането на мускулните влакна. Важна структурна и функционална формация на мускула е моторна или невромоторна единица, състояща се от един моторен неврон и мускулните влакна, инервирани от него. Различават се малки, средни и големи двигателни единици в зависимост от броя на мускулните влакна, участващи в акта на свиване.
Система от слоеве и мембрани на съединителната тъкан свързва мускулните влакна в една работна система, която с помощта на сухожилията предава тягата, която възниква по време на свиване на мускулите, към костите на скелета.
Целият мускул е пронизан от разклонена мрежа от кръвоносни съдове и лимфни разклонения. смукатели. Червени мускулни влакнаимат по-плътна мрежа от кръвоносни съдове от бяло.Те имат голям запас от гликоген и липиди, характеризират се със значителна тонизираща активност, способност да издържат на продължителен стрес и да извършват продължителна динамична работа. Всяко червено влакно има повече от белите митохондрии – генератори и доставчици на енергия, заобиколени от 3-5 капиляра, което създава условия за по-интензивно кръвоснабдяване на червените влакна и високо ниво на метаболитни процеси.
Бели мускулни влакнаимат миофибрили, които са по-дебели и по-здрави от миофибрилите на червените влакна; те се свиват бързо, но не са способни на продължително напрежение. Митохондриите на бялото вещество имат само един капиляр. Повечето мускули съдържат червени и бели влакна в различни пропорции. Има и мускулни влакна тоник(способен на локално възбуждане без разпространение); фаза,.способни да реагират на разпространяваща се вълна от възбуждане както със свиване, така и с отпускане; преходен, съчетаващ и двата свойства.
Мускулна помпа- физиологична концепция, свързана с мускулната функция и нейния ефект върху собственото кръвоснабдяване. Основното му действие се проявява, както следва: при свиване на скелетните мускули се забавя притока на артериална кръв към тях и се ускорява изтичането й през вените; в периода на релаксация венозният отток намалява, а артериалният приток достига своя максимум. Обменът на вещества между кръвта и тъканната течност се осъществява през капилярната стена.

Ориз. 2.5. Схематично представяне на процесите, протичащи в
синапс при възбуждане:
1 - синаптични везикули, 2 - пресинаптична мембрана, 3 - посредник, 4 - постсинаптична мембрана, 5 - синаптична цепнатина
Механизми на мускулитеМускулните функции се регулират от различни намаления отдели на централната нервна система (ЦНС), които до голяма степен определят характера на тяхната многостранна дейност
(фази на движение, тонично напрежение и др.). РецепториДвигателният апарат поражда аферентни влакна на двигателния анализатор, които съставляват 30-50% от влакната на смесените (аферентно-еферентни) нерви, насочени към гръбначния мозък. Мускулна контракция Предизвиква импулси, които са източник на мускулно усещане - кинестезия.
Прехвърлянето на възбуждане от нервните влакна към мускулните влакна става чрез нервно-мускулна връзка(Фиг. 2.5), която се състои от две мембрани, разделени с процеп – пресинаптична (с нервен произход) и постсинаптична (с мускулен произход). При излагане на нервен импулс се освобождават кванти ацетилхолин, което води до появата на електрически потенциал, който може да възбуди мускулните влакна. Скоростта на предаване на нервния импулс през синапса е хиляди пъти по-малка, отколкото в нервното влакно. Провежда възбуждането само по посока на мускула. Обикновено до 150 импулса могат да преминат през нервно-мускулната връзка на бозайниците за една секунда. При умора (или патология) подвижността на нервно-мускулните окончания намалява и естеството на импулсите може да се промени.
Химия и енергия на мускулната контракция.Свиването и напрежението на мускула се извършва поради енергията, освободена по време на химични трансформации, които се случват при навлизане в
мускул с нервен импулс или прилагане на директно дразнене към него. Химичните трансформации в мускулите се случват като в присъствието на кислород(при аеробни условия) и в негово отсъствие(при анаеробни условия).
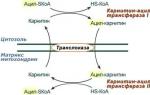
Разцепване и ресинтез на аденозин трифосфорна киселина (АТФ).Основният източник на енергия за мускулна контракция е разграждането на АТФ (намира се в клетъчната мембрана, ретикулума и миозиновите нишки) в аденозин дифосфорна киселина (ADP) и фосфорни киселини. В този случай 10 000 кал се освобождават от всеки грам молекула АТФ:
ATP = ADP + H3PO4 + 10 000 кал.
По време на по-нататъшни трансформации ADP се дефосфорилира до аденилова киселина. Разграждането на АТФ се стимулира от протеиновия ензим актомиозин (аденозин трифосфатаза). В покой не е активен, активира се при възбуда на мускулните влакна. На свой ред АТФ действа върху миозиновите нишки, увеличавайки тяхната разтегливост. Активността на актомиозина се увеличава под въздействието на Ca йони, които в покой се намират в саркоплазмения ретикулум.
Резервите на АТФ в мускулите са незначителни и за поддържане на тяхната активност е необходим непрекъснат ресинтез на АТФ. Това се дължи на енергията, получена от разграждането на креатин фосфат (CrP) до креатин (Cr) и фосфорна киселина (анаеробна фаза). С помощта на ензими фосфатната група от KrP бързо се прехвърля в ADP (в рамките на хилядни от секундата). В този случай за всеки мол CrP се отделят 46 kJ:
По този начин, последният процес, който осигурява целия енергиен разход на мускула, е процесът на окисление.Междувременно дългосрочната мускулна активност е възможна само ако има достатъчно снабдяване с кислород, тъй като Съдържанието на вещества, способни да освобождават енергия, постепенно намалява при анаеробни условия.Освен това се натрупва млечна киселина; изместването на реакцията към киселинната страна нарушава ензимните реакции и може да доведе до инхибиране и дезорганизация на метаболизма и намаляване на мускулната ефективност. Подобни състояния възникват в човешкото тяло по време на работа с максимална, субмаксимална и висока интензивност (мощност), например при бягане на къси и средни разстояния. Поради развитата хипоксия (липса на кислород) АТФ не се възстановява напълно, възниква така нареченият кислороден дълг и се натрупва млечна киселина.
Аеробен ресинтез на АТФ(синоними: окислително фосфорилиране, тъканно дишане) - 20 пъти по-ефективен от анаеробното генериране на енергия.Частта от млечната киселина, натрупана по време на анаеробна дейност и в процеса на продължителна работа, се окислява до въглероден диоксид и вода (1/4-1/6 от него), получената енергия се използва за възстановяване на останалите части от млечна киселина в глюкоза и гликоген, като същевременно осигурява ресинтеза на АТФ и KrF. Енергията на окислителните процеси се използва и за ресинтеза на въглехидрати, необходими на мускула за неговата непосредствена дейност.
Като цяло въглехидратите осигуряват най-голямо количество енергия за мускулна работа. Например, по време на аеробното окисляване на глюкозата се образуват 38 молекули АТФ (за сравнение: по време на анаеробното разграждане на въглехидратите се образуват само 2 молекули АТФ).
Време за разгръщане на аеробния пътОбразуването на АТФ е 3-4 минути (за тренирани хора - до 1 минута), максималната мощност е 350-450 кал/мин/кг, времето за поддържане на максимална мощност е десетки минути. Ако в покой скоростта на аеробния ресинтез на АТФ е ниска, тогава по време на физическа активност неговата мощност става максимална и в същото време аеробният път може да работи с часове. Освен това е много икономичен: по време на този процес има дълбоко разлагане на изходните вещества до крайните продукти CO2 и NaO. В допълнение, аеробният път на ресинтеза на АТФ се отличава със своята гъвкавост при използването на субстрати: всички органични вещества в тялото се окисляват (аминокиселини, протеини, въглехидрати, мастни киселини, кетонови тела и др.).
Аеробният метод за ресинтез на АТФ обаче има и недостатъци: 1) той изисква консумация на кислород, чиято доставка до мускулната тъкан се осигурява от дихателната и сърдечно-съдовата система, което естествено е свързано с тяхното напрежение; 2) всякакви фактори, влияещи върху състоянието и свойствата на митохондриалните мембрани, нарушават образуването на АТФ; 3) развитието на аеробното образуване на АТФ е продължително във времето и с ниска мощност.
Мускулната активност, извършвана в повечето спортове, не може да бъде напълно осигурена от аеробния процес на ресинтез на АТФ и тялото е принудено да включва допълнително анаеробни методи за образуване на АТФ, които имат по-кратко време за разгръщане и по-голяма максимална мощност на процеса ( т.е. най-голямото количество АТФ, "образувано за единица време) - 1 мол АТФ съответства на 7,3 cal, или 40 J (1 cal == 4,19 J).
Връщайки се към анаеробните процеси на образуване на енергия, трябва да се изясни, че те протичат в най-малко два вида реакции: 1. Креатин фосфокиназа -когато CrP се разцепва, фосфорните групи от него се прехвърлят към ADP, като по този начин отново се синтезира ATP. Но запасите от креатин фосфат в мускулите са малки и това води до бързо (в рамките на 2-4 s) изчезване на този тип реакция. 2. Гликолитичен(гликолиза) - развива се по-бавно, в рамките на 2-3 минути интензивна работа. Гликолизата започва с фосфорилирането на мускулния гликоген и кръвната глюкоза. Енергията на този процес е достатъчна за няколко минути усилена работа. На този етап завършва първият етап на фосфорилиране на гликоген и се извършва подготовка за окислителния процес. След това идва вторият етап от гликолитичната реакция - дехидрогениране и третият - редукцията на АДФ до АТФ. Гликолитичната реакция завършва с образуването на две молекули млечна киселина, след което се развиват дихателни процеси (на 3-5 минути работа), когато млечната киселина (лактат), образувана по време на анаеробни реакции, започва да се окислява.
Биохимични показатели за оценка на креатин фосфат анаеробния път на ресинтеза на АТФ са креатининовият коефициент и алактатният (без млечна киселина) кислороден дълг. Креатининово съотношение- е екскрецията на креатинин в урината на ден на 1 kg телесно тегло. При мъжете екскрецията на креатинин варира от 18-32 mg/ден x kg, а при жените – 10-25 mg/ден x kg. Съществува линейна зависимост между съдържанието на креатин фосфат и образуването на креатинин. Следователно, като се използва креатининовият коефициент, могат да се оценят потенциалните възможности на този път на ресинтеза на АТФ.
Биохимични промени в организма, причинени от натрупването на млечна киселинав резултат на гликолиза. Ако е в покой преди началото на цервикалната активност концентрация на лактатв кръвта е 1-2 mmol/l, то след интензивно, краткотрайно натоварване за 2-3 минути тази стойност може да достигне 18-20 mmol/l. Друг показател, отразяващ натрупването на млечна киселина в кръвта е кръвна картина(pH): в покой 7,36, след тренировка намалява до 7,0 или повече. Натрупването на лактат в кръвта определя неговата алкален резерв -алкални компоненти на всички кръвни буферни системи.
Краят на интензивната мускулна дейност е придружен от намаляване на кислородната консумация - първоначално рязко, след това по-плавно. В тази връзка подчертават два компонента на кислородния дълг:бърз (алактат) и бавен (лактат). лактат -това е количеството кислород, което се използва след приключване на работата за елиминиране на млечна киселина: по-малка част се окислява до J-bO и COa, по-голямата част се превръща в гликоген. Тази трансформация изисква значително количество АТФ, което се образува аеробно благодарение на кислорода, който представлява лактатен дълг.Метаболизмът на лактат се осъществява в клетките на черния дроб и миокарда.
Количеството кислород, необходимо за пълно осигуряване на извършената работа, се нарича потребност от кислород.Например, при състезание на 400 м, потребността от кислород е приблизително 27 литра. Времето за изминаване на разстоянието на ниво световен рекорд е около 40 секунди. Проучванията показват, че през това време спортистът усвоява 3-4 литра 02. Следователно 24 литра са общ кислороден дълг(около 90% от потреблението на кислород), което се елиминира след състезанието.
В състезанието на 100 м кислородният дефицит може да достигне до 96% от търсенето. При бягане на 800 м делът на анаеробните реакции леко намалява - до 77%, при бягане на 10 000 м - до 10%, т.е. преобладаващата част от енергията се доставя чрез дихателни (аеробни) реакции.
Механизмът на мускулна релаксация.Веднага след като нервните импулси престанат да навлизат в мускулните влакна, Ca2 йони, под действието на така наречената калциева помпа, поради енергията на АТФ, отиват в цистерните на саркоплазмения ретикулум и концентрацията им в саркоплазмата намалява до оригинала. ниво. Това причинява промени в конформацията на тропонина, който чрез фиксиране на тропомиозин в определена област на актинови нишки прави невъзможно образуването на напречни мостове между дебели и тънки нишки. Благодарение на еластичните сили, които възникват по време на мускулна контракция в колагеновите нишки, обграждащи мускулното влакно, то се връща в първоначалното си състояние при отпускане. По този начин процесът на мускулна релаксация или релаксация, както и процесът на мускулна контракция, се осъществяват с помощта на енергията на хидролизата на АТФ.
По време на мускулната активност процесите на свиване и отпускане протичат последователно в мускулите и следователно скоростно-силовите качества на мускулите еднакво зависят от скоростта на мускулното съкращение и от способността на мускулите да се отпуснат.
Кратка характеристика на гладкомускулните влакна.Гладките мускулни влакна нямат миофибрили. Тънките нишки (актин) са свързани със сарколемата, дебелите нишки (миозин) са разположени вътре в мускулните клетки. Гладките мускулни влакна също нямат цистерни с Ca йони. Под въздействието на нервен импулс Ca йони бавно навлизат в саркоплазмата от извънклетъчната течност и също бавно напускат, след като нервните импулси спрат да пристигат. Следователно гладките мускулни влакна се свиват бавно и се отпускат бавно.
Общ преглед на скелета човешки мускули.Мускули на багажника(фиг. 2.6 и 2.7) включват мускулите на гръдния кош, гърба и корема. Мускулите на гръдния кош участват в движенията на горните крайници, а също така осигуряват доброволни и неволни дихателни движения. Дихателните мускули на гръдния кош се наричат външни и вътрешни междуребрени мускули. Дихателната мускулатура включва и диафрагмата. Мускулите на гърба се състоят от повърхностни и дълбоки мускули. Повърхностните осигуряват някои движения на горните крайници, главата и шията. Дълбоките („изправители на багажника“) са прикрепени към спинозните процеси на прешлените и се простират по гръбначния стълб. Мускулите на гърба участват в поддържането на вертикалното положение на тялото; при силно напрежение (свиване) те карат тялото да се огъва назад. Коремните мускули поддържат налягането в коремната кухина (корема), участват в някои движения на тялото (огъване на торса напред, огъване и завъртане на страни), както и по време на процеса на дишане.
Мускулите на главата и шията -мимически, дъвчене и движение на главата и шията. Лицевите мускули са прикрепени в единия край към костта, в другия към кожата на лицето, някои могат да започват и завършват в кожата. Мускулите на лицето осигуряват движенията на кожата на лицето, отразяват различни психични състояния на човек, придружават речта и са важни в комуникацията. Когато дъвкателните мускули се свиват, те карат долната челюст да се движи напред и настрани. Мускулите на врата участват в движенията на главата. Задната група мускули, включително мускулите на гърба на главата, с тонично (от думата „тон“) свиване държи главата в изправено положение.

Ориз. 2.6. Мускули на предната половина на тялото (според Силванович):
1 - темпорален мускул, 2 - дъвкателен мускул, 3 - стерноклеидомастоиден мускул, 4 - голям гръден мускул, 5 - среден скален мускул, b - външен наклонен мускул на корема, 7 - vastus medialis, 8 - vastus lateralis, 9 - прав бедрен мускул, 10 - сарториус, 11 - нежен мускул 12 - вътрешен наклонен коремен мускул, 13 - прав коремен мускул, 14 - бицепс brachii мускул, 15 ~ външни междуребрени мускули, 16 - orbicularis oris мускул, 17 - orbicularis oculi мускул, 18 - фронтален мускул
Мускули на горните крайнициосигуряват движение на раменния пояс, рамото, предмишницата и движат ръката и пръстите. Основните мускули-антагонисти са бицепсите (флексорите) и трицепсите (екстензорите) на рамото. Движенията на горния крайник и преди всичко на ръката са изключително разнообразни. Това се дължи на факта, че ръката служи като човешки орган на труда.

Ориз. 2.7. Мускули на задната половина на тялото (според Силванович):
1 - ромбоиден мускул, 2 - токоизправител на торса, 3 - дълбоки мускули на глутеалния мускул, 4 - двуглав бедрен мускул, 5 - телешки мускул, 6 - ахилесово сухожилие, 7 - седалищен мускул, 8 - latissimus skipae мускул, 9 - делтоид, 10 - трапецовиден мускул
Мускулите на долните крайнициосигуряват движение на бедрото, подбедрицата и стъпалото. Бедрените мускули играят важна роля за поддържане на изправено положение на тялото, но при хората те са по-развити, отколкото при другите гръбначни животни. Мускулите, които извършват движения на долната част на крака, са разположени на бедрото (например четириглавият мускул, чиято функция е да разтяга долния крак в колянната става; антагонистът на този мускул е мускулът на бицепса на бедрената кост). Стъпалото и пръстите на краката се задвижват от мускули, разположени в подбедрицата и стъпалото. Огъването на пръстите на краката се осъществява чрез свиване на мускулите, разположени на ходилото, и удължаване от мускулите на предната повърхност на крака и стъпалото. Много мускули на бедрото, крака и стъпалото участват в поддържането на човешкото тяло в изправено положение.
СЪСструктура и свиване на мускулните влакна.
Мускулната контракция в живата система е механохимичен процес. Съвременната наука го смята за най-съвършената форма на биологична мобилност. Биологичните обекти „разработиха“ свиването на мускулните влакна като начин за придвижване в пространството (което значително разшири техните жизнени възможности).
Мускулната контракция се предшества от фаза на напрежение, която е резултат от работата, извършена чрез преобразуване на химическата енергия в механична енергия директно и с добра ефективност (30-50%). Натрупването на потенциална енергия във фазата на напрежение привежда мускула в състояние на възможно, но все още неосъществено свиване.
Животните и хората имат (и хората вярват, че вече са добре проучени) два основни вида мускули:набразден и гладък. Набраздени мускулиили скелетни са прикрепени към костите (с изключение на набраздените влакна на сърдечния мускул, които се различават от скелетните мускули по състав). Гладка мускулиподдържат тъканите на вътрешните органи и кожата и формират мускулите на стените на кръвоносните съдове, както и на червата.
В биохимията на спорта изучават скелетни мускули, „конкретно отговорен“ за спортните резултати.
Мускулът (като макроформация, принадлежаща на макрообект) се състои от отделни мускулни влакна(микро образувания). Има хиляди от тях в мускула; съответно мускулното усилие е интегрална стойност, която обобщава контракциите на много отделни влакна. Има три вида мускулни влакна: бялобързо потрепващ , междиненИ червенбавно потрепващ. Видовете влакна се различават по механизма на тяхното енергоснабдяване и се контролират от различни моторни неврони. Типовете мускули се различават по отношение на видовете влакна.
Отделно мускулно влакно - нишковидно ацелуларно образувание - симпласт. Симпластът „не прилича на клетка“: има силно удължена форма с дължина от 0,1 до 2-3 cm, в сарториусния мускул до 12 cm и дебелина от 0,01 до 0,2 mm. Симпластът е заобиколен от черупка - сарколема,към чиято повърхност се приближават окончанията на няколко двигателни нерва. Sarcolemma е двуслойна липопротеинова мембрана (с дебелина 10 nm), подсилена от мрежа от колагенови влакна. Когато се отпуснат след контракция, те връщат симпласта в първоначалната му форма (фиг. 4).
Ориз. 4. Индивидуално мускулно влакно.
На външната повърхност на мембраната на сарколема винаги се поддържа електрически мембранен потенциал, дори в покой е равен на 90-100 mV. Наличието на потенциал е необходимо условие за контролиране на мускулните влакна (като автомобилен акумулатор). Потенциалът се създава благодарение на активния (тоест с разход на енергия - АТФ) пренос на вещества през мембраната и нейната селективна пропускливост (според принципа - "който искам, ще го пусна или ще го пусна" ). Следователно вътре в симпласта някои йони и молекули се натрупват в по-високи концентрации, отколкото навън.
Сарколемата е добре пропусклива за K + йони - те се натрупват вътре, а Na + йони се отстраняват навън. Съответно концентрацията на Na + йони в междуклетъчната течност е по-голяма от концентрацията на K + йони вътре в симпласта. Изместването на pH към киселинната страна (например по време на образуването на млечна киселина) повишава пропускливостта на сарколемата за високомолекулни вещества (мастни киселини, протеини, полизахариди), които обикновено не преминават през нея. Веществата с ниско молекулно тегло (глюкоза, млечна и пирогроздена киселина, кетонови тела, аминокиселини, къси пептиди) лесно преминават (дифундират) през мембраната.
Вътрешно съдържание на simplast – саркоплазма– Това е колоидна протеинова структура (консистенцията наподобява желе). В суспендирано състояние съдържа гликогенни включвания, мастни капчици и различни субклетъчни частици са „вградени“: ядра, митохондрии, миофибрили, рибозоми и др.
Контрактилен „механизъм“ вътре в симпласта – миофибрили.Това са тънки (Ø 1 - 2 микрона) мускулни нишки, дълги - почти равни на дължината на мускулното влакно. Установено е, че в симпластите на нетренираните мускули миофибрилите не са разположени подредено по симпласта, а с разсейки и отклонения, а в тренираните миофибрилите са ориентирани по надлъжната ос и също са групирани в снопове, като въжета. (При предене на изкуствени и синтетични влакна макромолекулите на полимера първоначално не са разположени стриктно по протежение на влакното и, подобно на спортистите, те се „упорито тренират“ - ориентирани правилно - по оста на влакната, чрез многократно пренавиване: вижте дългия работилници на ЗИВ и Химволокно).
Под светлинен микроскоп може да се види, че миофибрилите наистина са „набраздени“. Те редуват светли и тъмни зони - дискове. Тъмни джанти А (анизотропни) протеини съдържат повече от леки дискове аз (изотропен). Леки дискове, пресечени от мембрани З (телофрагми) и участък от миофибрила между две З - наречени мембрани саркомер. Миофибрилата се състои от 1000 – 1200 саркомера (фиг. 5).
Съкращението на мускулното влакно като цяло се състои от отделни контракции саркомери.Свивайки се всеки поотделно, саркомерите заедно създават интегрална сила и извършват механична работа за свиване на мускула.
Дължината на саркомера варира от 1,8 µm в покой до 1,5 µm по време на умерено и до 1 µm по време на пълна контракция. Дисковете на саркомерите, тъмни и светли, съдържат протофибрили (миофиламенти) - белтъчни нишковидни структури. Те се срещат в два вида: дебели (Ø – 11 – 14 nm, дължина – 1500 nm) и тънки (Ø – 4 – 6 nm, дължина – 1000 nm).

Ориз. 5. Миофибрилна зона.
Леки колела ( аз ) се състоят само от тънки протофибрили и тъмни дискове ( А ) – от протофибрили от два вида: тънки, закрепени заедно с мембрана, и дебели, концентрирани в отделна зона ( з ).
Когато саркомерът се свие, дължината на тъмния диск ( А ) не се променя и дължината на светлинния диск ( аз ) намалява, когато тънките протофибрили (светли дискове) се придвижват в пространствата между дебелите (тъмните дискове). На повърхността на протофибрилите има специални израстъци - сраствания (с дебелина около 3 nm). В “работно положение” те образуват зацепване (напречни мостове) между дебели и тънки нишки от протофибрили (фиг. 6). При договаряне З -мембраните опират в краищата на дебелите протофибрили, а тънките протофибрили могат дори да се увият около дебелите. По време на суперконтракцията краищата на тънките нишки в центъра на саркомера се навиват и краищата на дебелите протофибрили се смачкват.

Ориз. 6. Образуване на сраствания между актин и миозин.
Енергийното снабдяване на мускулните влакна се осъществява с помощта на саркоплазмен ретикулум(известен още като - саркоплазмен ретикулум) – системи от надлъжни и напречни тръби, мембрани, мехурчета, отделения.
В саркоплазмения ретикулум протичат различни биохимични процеси по организиран и контролиран начин, мрежата обхваща всичко заедно и всяка миофибрила поотделно. Ретикулумът включва рибозоми, те извършват синтеза на протеини и митохондрии - „клетъчни енергийни станции“ (както е определено в училищния учебник). Всъщност митохондриитевградени между миофибрилите, което създава оптимални условия за енергийно захранване на процеса на мускулна контракция. Установено е, че в тренираните мускули броят на митохондриите е по-голям, отколкото в същите нетренирани мускули.
Химичен състав на мускулите.
Вода съсоставя 70 - 80% от мускулното тегло.
катерици. Протеините представляват от 17 до 21% от мускулното тегло: приблизително 40% от всички мускулни протеини са концентрирани в миофибрилите, 30% в саркоплазмата, 14% в митохондриите, 15% в сарколемата, останалите в ядрата и други клетъчни органели.
Мускулната тъкан съдържа ензимни миогенни протеинигрупи, миоалбумин– резервен протеин (съдържанието му постепенно намалява с възрастта), червен протеин миоглобин– хромопротеин (нарича се мускулен хемоглобин, свързва повече кислород от кръвния хемоглобин), а също глобулини, миофибриларни протеини.Повече от половината от миофибриларните протеини са миозин, около една четвърт - актин, останалото е тропомиозин, тропонин, α- и β-актинини, ензими креатин фосфокиназа, деаминаза и др. Мускулната тъкан съдържа ядренкатерици– нуклеопротеини, митохондриални протеини.В протеини строма,сплитане на мускулна тъкан - основната част - колагенИ еластинсарколеми, както и миостромини (свързани с З -мембрани).
впредварително разтворими азотни съединения.Човешките скелетни мускули съдържат различни водоразтворими азотни съединения: АТФ,от 0,25 до 0,4%, креатин фосфат (CrP)– от 0,4 до 1% (с тренировка количеството му се увеличава), разпадните им продукти са ADP, AMP, креатин. В допълнение, мускулите съдържат дипептид карнозин,около 0,1 - 0,3%, участващи във възстановяването на работата на мускулите по време на умора; карнитин,отговорен за транспортирането на мастни киселини през клетъчните мембрани; аминокиселини, а сред тях преобладава глутаминът (това обяснява ли използването на мононатриев глутамат, прочетете състава на подправките, за да придадете на храната вкус на месо); пуринови бази, урея и амоняк. Скелетните мускули също съдържат около 1,5% фосфатиди,които участват в тъканното дишане.
Без азот връзки. Мускулите съдържат въглехидрати, гликоген и неговите метаболитни продукти, както и мазнини, холестерол, кетонови тела и минерални соли. В зависимост от диетата и степента на тренировка, количеството гликоген варира от 0,2 до 3%, докато тренировките увеличават масата на свободния гликоген. Мазнините за съхранение се натрупват в мускулите по време на тренировка за издръжливост. Свързаните с протеин мазнини съставляват приблизително 1%, а мембраните на мускулните влакна могат да съдържат до 0,2% холестерол.
Минерали.Минералите в мускулната тъкан съставляват приблизително 1 - 1,5% от мускулното тегло; това са главно калиеви, натриеви, калциеви и магнезиеви соли. Минералните йони като K +, Na +, Mg 2+, Ca 2+, Cl -, HP0 4 ~ играят жизненоважна роля в биохимичните процеси по време на мускулна контракция (те са включени в „спортните“ добавки и минералната вода).
Биохимия на мускулните протеини.
Основният контрактилен протеин на мускулите е миозинсе отнася до фибриларни протеини (молекулно тегло около 470 000). Важна характеристика на миозина е способността да образува комплекси с ATP и ADP молекули (което ви позволява да „вземете“ енергия от ATP) и с протеина актин (което прави възможно поддържането на контракция).
Молекулата на миозина има отрицателен заряд и специфично взаимодейства с Ca ++ и Mg ++ йони. Миозинът, в присъствието на Са++ йони, ускорява хидролизата на АТФ и по този начин проявява ензимни активност на аденозин трифосфат:
миозин-АТФ+H2O → миозин + ADP + H3PO4 + работа(енергия 40 kJ/mol)
Миозиновият протеин се образува от две еднакви, дълги полипептидни α-вериги, усукани като двойна спирала, фиг.7. Под действието на протеолитичните ензими молекулата на миозина се разпада на две части. Една от неговите части е способна да се свързва с актин чрез сраствания, образувайки актомиозин. Тази част е отговорна за активността на аденозинтрифосфатазата, която зависи от рН на околната среда, оптималното е рН 6,0 - 9,5, както и концентрацията на KCl. Актомиозиновият комплекс се разпада в присъствието на АТФ, но при липса на свободен АТФ е стабилен. Втората част на миозиновата молекула също се състои от две усукани спирали; поради електростатичен заряд те свързват миозиновите молекули в протофибрили.

Ориз. 7. Строеж на актомиозин.
Вторият по важност контрактилен протеин е актин(фиг. 7). Може да съществува в три форми: мономерна (глобуларна), димерна (глобуларна) и полимерна (фибриларна). Мономерният глобуларен актин, когато неговите полипептидни вериги са плътно опаковани в компактна сферична структура, се свързва с АТФ. Чрез разделяне на ATP, актиновите мономери - A, образуват димери, включително ADP: A - ADP - A. Полимерният фибриларен актин е двойна спирала, състояща се от димери, Фиг. 7.
Глобуларният актин се трансформира във фибриларен актин в присъствието на K + и Mg ++ йони, а фибриларният актин преобладава в живите мускули.
Миофибрилите съдържат значително количество протеин тропомиозин, който се състои от две α-спирални полипептидни вериги. В почиващите мускули той образува комплекс с актин и блокира неговите активни центрове, тъй като актинът е в състояние да се свърже с Ca ++ йони, които премахват тази блокада.
На молекулярно ниво дебелите и тънките протофибрили на саркомера взаимодействат електростатично, тъй като имат специални области - израстъци и издатини - където се образува заряд. В областта на А-диска дебелите протофибрили са изградени от сноп от надлъжно ориентирани миозинови молекули, тънките протофибрили са подредени радиално около дебелите, образувайки структура, подобна на многожилен кабел. В централната М-лента на дебели протофибрили миозиновите молекули са свързани с техните „опашки“, а техните изпъкнали „глави“ - израстъци са насочени в различни посоки и са разположени по редовни спирални линии. Всъщност срещу тях във фибриларните актинови спирали на известно разстояние една от друга също изпъкват мономерни актинови глобули. Всяка издатина има активен център,поради което е възможно образуването на сраствания с миозин. Z-мембраните на саркомерите (като редуващи се пиедестали) държат тънки протофибрили заедно.
Биохимия на контракцията и релаксацията.
Цикличните биохимични реакции, които се случват в мускула по време на свиване, осигуряват многократно образуване и разрушаване на срастванията между „главите“ - израстъците на миозиновите молекули на дебелите протофибрили и издатините - активните центрове на тънките протофибрили. Работата по образуване на сраствания и придвижване на актиновата нишка по миозиновата нишка изисква както прецизен контрол, така и значителен разход на енергия. Реално в момента на свиване на влакната се образуват около 300 сраствания в минута във всеки активен център - протрузия.
Както отбелязахме по-рано, само ATP енергията може директно да се преобразува в механична работа на мускулна контракция. АТФ, хидролизиран от ензимния център на миозина, образува комплекс с целия миозинов протеин. В комплекса АТФ-миозин, миозинът, наситен с енергия, променя структурата си, а с това и външните "размери" и по този начин извършва механична работа за съкращаване на растежа на миозиновата нишка.
В покойния мускул миозинът все още е свързан с АТФ, но чрез Mg++ йони без хидролитично разцепване на АТФ. Образуването на сраствания между миозина и актина в покой се предотвратява от комплекса на тропомиозин с тропонин, който блокира активните центрове на актина. Блокадата се поддържа и АТФ не се разгражда, докато Ca++ йоните се свързват. Когато нервен импулс пристигне в мускулно влакно, той се освобождава импулсен предавател– неврохормон ацетилхолин. Na+ йоните неутрализират отрицателния заряд на вътрешната повърхност на сарколемата и я деполяризират. В този случай Ca++ йони се освобождават и се свързват с тропонина. На свой ред тропонинът губи заряда си, което води до деблокиране на активните центрове - издатините на актиновите нишки - и възникват сраствания между актина и миозина (тъй като електростатичното отблъскване на тънки и дебели протофибрили вече е премахнато). Сега, в присъствието на Ca ++, ATP взаимодейства с центъра на ензимната активност на миозина и се разцепва, а енергията на трансформиращия комплекс се използва за намаляване на адхезията. Веригата от молекулярни събития, описани по-горе, е подобна на електрически ток, презареждащ микрокондензатор; неговата електрическа енергия веднага се преобразува в механична работа на място и трябва да се презареди отново (ако искате да продължите).
След разкъсването на адхезива АТФ не се разцепва, а отново образува ензимно-субстратен комплекс с миозин:
M–A + ATP -----> M – ATP + Aили
M–ADP–A + ATP ----> M–ATP + A + ADP
Ако в този момент пристигне нов нервен импулс, тогава реакциите на „презареждане“ се повтарят; ако следващият импулс не пристигне, мускулът се отпуска. Връщането на свит мускул при отпускане в първоначалното му състояние се осигурява от еластичните сили на протеините в мускулната строма. Излагайки съвременни хипотези за свиване на мускулите, учените предполагат, че в момента на свиване актиновите нишки се плъзгат по миозиновите нишки и тяхното скъсяване също е възможно поради промени в пространствената структура на контрактилните протеини (промени във формата на спиралата).
В покой АТФ има пластифициращ ефект: като се комбинира с миозин, той предотвратява образуването на адхезии с актин. Разграждайки се по време на мускулна контракция, АТФ осигурява енергия за процеса на скъсяване на срастванията, както и за работата на „калциевата помпа“ - доставката на Ca ++ йони. Разграждането на АТФ в мускулите става с много висока скорост: до 10 микромола на 1 g мускул на минута. Тъй като общите резерви на АТФ в мускула са малки (може да са достатъчни само за 0,5-1 секунда работа при максимална мощност), за да се осигури нормална мускулна активност, АТФ трябва да се възстановява със същата скорост, с която се разгражда.