stellate cells. Could perisinusoidal cells be regional liver stem cells? Clinical liver cytology: Ito stellate cells
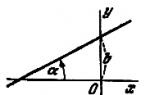
Top - Schematic representation of the Ito cell (HSC) in the neighborhood of the nearest hepatocytes (PC), below the sinusoidal liver epithelial cells (EC). S - liver sinusoid; KC - Kupffer cell. Bottom left - Ito cells in culture under a light microscope. Bottom right - Electron microscopy reveals numerous fat vacuoles (L) of Ito cells (HSCs) that store retinoids.
Ito cells(synonyms: stellate cell of the liver, fat storage cell, lipocyte, English Hepatic Stellat Cell, HSC, Cell of Ito, Ito cell) - pericytes contained in, able to function in two different states - calm and activated. Activated Ito cells play a major role in the formation of scar tissue in liver damage.
In an intact liver, stellate cells are found in calm state. In this state, the cells have several outgrowths that surround the sinusoidal capillary. Another distinguishing feature of cells is the presence in their cytoplasm of reserves of vitamin A (retinoid) in the form of fat droplets. Quiet Ito cells make up 5-8% of all liver cells.
Outgrowths of Ito cells are divided into two types: perisinusoidal(subendothelial) and interhepatocellular. The first ones leave the cell body and extend along the surface of the sinusoidal capillary, covering it with thin finger-shaped branches. Perisinusoidal outgrowths are covered with short villi and have characteristic long microprotrusions extending even further along the surface of the capillary endothelial tube. Interhepatocellular outgrowths, having overcome the plate of hepatocytes and reaching the neighboring sinusoid, are divided into several perisinusoidal outgrowths. Thus, the Ito cell covers, on average, slightly more than two adjacent sinusoids.
When the liver is damaged, Ito cells become activated state. The activated phenotype is characterized by proliferation, chemotaxis, contractility, loss of retinoid stores, and production of myofibroblastic-like cells. Activated liver stellate cells also show increased levels of new genes such as ICAM-1, chemokines and cytokines. Activation indicates the beginning of an early stage of fibrogenesis and precedes the increased production of ECM proteins. The final stage of liver healing is characterized by increased apoptosis of activated Ito cells, as a result of which their number is sharply reduced.
Gold chloride staining is used to visualize Ito cells under microscopy. It has also been established that a reliable marker for the differentiation of these cells from other myofibroblasts is their expression of the reelin protein.
Story [ | ]
In 1876 Karl von Kupfer described the cells he named "Sternzellen" (stellate cells). When stained with gold oxide, inclusions were visible in the cytoplasm of the cells. Mistakenly considering them to be fragments of erythrocytes captured by phagocytosis, Kupfer in 1898 revised his views on the "stellate cell" as a separate type of cell and classified them as phagocytes. However, in subsequent years, descriptions of cells similar to Kupffer's "stellate cells" appeared regularly. They were given various names: interstitial cells, parasinusoid cells, lipocytes, pericytes. The role of these cells remained a mystery for 75 years, until a professor (Toshio Ito) discovered some cells containing blotches of fat in the perisinusoidal space of the human liver. Ito called them "shibo-sesshu saibo" - fat-absorbing cells. Realizing that the inclusions were fat produced by cells from glycogen, he changed the name to "shibo-chozo saibo" - fat-storing cells. AT
Keywords
LIVER / ITO STAR CELLS/ MORPHOLOGY / CHARACTERISTIC / VITAMIN A / FIBROSISannotation scientific article on fundamental medicine, author of scientific work - Tsyrkunov V.M., Andreev V.P., Kravchuk R.I., Kondratovich I.A.
Introduction. The role of Ito stellate cells (ISCs) is defined as one of the leading ones in the development of fibrosis in the liver, however, in vivo visualization of the structure of ITOs in clinical practice is used minimally. The purpose of the work: to present the structural and functional characteristics of HCI based on the results of cytological identification of intravital liver biopsy specimens. Materials and methods. Classical methods of light and electron microscopy of biopsy specimens and original techniques using ultrathin sections, fixation and staining were used. Results. Photo-illustrations of light and electron microscopy of liver biopsy specimens from patients with chronic hepatitis C show the structural characteristics of HSCs at different stages (rest, activation) and in the process of transformation into myofibroblasts. Conclusions. The use of original methods of clinical morphological identification and assessment of the functional state of HCI will improve the quality of diagnosis and prediction of liver fibrosis.
Related Topics scientific works on fundamental medicine, the author of scientific work - Tsyrkunov V.M., Andreev V.P., Kravchuk R.I., Kondratovich I.A.
-
Clinical Liver Cytology: Kupffer cells
2017 / Tsyrkunov V.M., Andreev V.P., Kravchuk R.I., Prokopchik N.I. -
Monitoring the morphological effects of autologous mesenchymal stem cells transplanted into the liver in viral cirrhosis (clinical observation)
2018 / Aukashnk S.P., Alenikova O.V., Tsyrkunov V.M., Isaykina Ya.I., Kravchuk R.I. -
Clinical morphology of the liver: necrosis
2017 / Tsyrkunov V.M., Prokopchik N.I., Andreev V.P., Kravchuk R.I. -
Polymorphism of liver stellate cells and their role in fibrogenesis
2008 / Aidagulova S.V., Kapustina V.I. -
The structure of sinusoidal liver cells in patients with co-infection with HIV / hepatitis C virus
2013 / Matievskaya N. V., Tsyrkunov V. M., Kravchuk R. I., Andreev V. P. -
Mesenchymal stem cells as a promising method for the treatment of liver fibrosis/cirrhosis
2013 / Lukashik S. P., Aleinikova O. V., Tsyrkunov V. M., Isaykina Ya. I., Romanova O. N., Shimansky A. T., Kravchuk R. I. -
Isolation and cultivation of rat liver myofibroblasts by explantation
2012 / Miyanovich O., Shafigullina A. K., Rizvanov A. A., Kiyasov A. P. -
Pathological aspects of the formation of liver fibrosis in HCV infection and other liver lesions: modern concepts
2009 / Lukashik S. P., Tsyrkunov V. M. -
Analysis of rat myofibroblasts obtained from the structures of the portal tracts of the liver by explantation
2013 / Miyanovich O., Katina M. N., Rizvanov A. A., Kiyasov A. P. -
Transplanted liver stellate cells are involved in organ regeneration after partial hepatectomy without the risk of developing liver fibrosis
2012 / Shafigullina A. K., Gumerova A. A., Trondin A. A., Titova M. A., Gazizov I. M., Burganova G. R., Kaligin M. S., Andreeva D. I., Rizvanov A. A., Mukhammedov A. R., Kiyasov A. P.
introduction. The role of Ito stellate cells (Hepatic Stellate Cells , HSC) has been identified as one of the leading in the development of liver fibrosis , but the use of intravital visualization of HSC structures in clinical practice is minimal. The aim of the work is to present the structural and functional characteristic of HSC based on the findings of cytological identification of intravital liver biopsy samples. materials and methods. Classical methods of light and electron microscopy of biopsy samples within the original technique of using ultrathin sections, fixation and staining were applied. results. The structural characteristics of the HSC of the liver biopsy samples from patients with chronic hepatitis C are presented on photo illustrations of light and electron microscopy. HSCs are depicted at different stages (rest, activation) and during the process of transformation into myofibroblasts. Conclusions. The use of original methods of clinical and morphological identification and evaluation of the functional status of HSC allows to improve the quality of diagnosis and prognosis of liver fibrosis.
The text of the scientific work on the topic "Clinical Liver Cytology: Ito stellate cells"
UDK 616.36-076.5
CLINICAL LIVER CYTOLOGY: ITO Stellate Cells
Tsyrkunov V. M. ( [email protected]), Andreev V.P. ( [email protected]), Kravchuk R. I. ( [email protected]), Kondratovich I. A. ( [email protected]) EE "Grodno State Medical University", Grodno, Belarus
Introduction. The role of Ito stellate cells (ISCs) is defined as one of the leading ones in the development of fibrosis in the liver, however, intravital visualization of the structure of ITOs in clinical practice is used minimally.
Purpose of the work: to present the structural and functional characteristics of HCI based on the results of cytological identification of intravital liver biopsy specimens.
Materials and methods. Classical methods of light and electron microscopy of biopsy specimens and original techniques using ultrathin sections, fixation and staining were used.
Results. Photo-illustrations of light and electron microscopy of liver biopsy specimens from patients with chronic hepatitis C show the structural characteristics of HSCs at different stages (rest, activation) and in the process of transformation into myofibroblasts.
Conclusions. The use of original methods for clinical morphological identification and assessment of the functional state of HCI will improve the quality of diagnosis and prediction of liver fibrosis.
Key words: liver, Ito stellate cells, morphology, characteristics, vitamin A, fibrosis.
Introduction
An unfavorable outcome of most chronic diffuse liver lesions of various etiologies, including chronic hepatitis C (CHC), is liver fibrosis, in the development of which the main participants are activated fibroblasts, the main source of which are activated Ito stellate cells (SSCs).
HSC, synonym - liver stellate cells, fat-storing cells, perisinusoidal lipocytes, stellate cells (English Hepatic Stellate Cell, HSC, Cell of Ito, Ito cell). ZKI were first described in 1876 by K. Kupffer and named by him stellate cells (“Stemzellen”). T. Ito, having found drops of fat in them, designated them at first fat-absorbing (“shibo-sesshusaibo”), and then, having established that fat was produced by the cells themselves from glycogen, fat-storing cells (“shibo-chozosaibo”) . In 1971, K. Wake proved the identity of Kupffer stellate cells and fat-storing Ito cells and that these cells “storage” vitamin A.
About 80% of vitamin A in the body accumulates in the liver, and up to 80% of all liver retinoids are deposited in HKI fatty drops. Retinol esters in chylomicrons enter hepatocytes, where they are converted to retinol, forming a complex of vitamin A with retinol-binding protein (RBP), which is secreted into the perisinusoidal space, from where it is deposited by cells.
Established by K. Popper, the close relationship between HCI and liver fibrosis demonstrated their dynamic rather than static function - the ability to directly participate in the remodeling of the intralobular perihepatocellular matrix.
The main method of morphological examination of the liver, which is carried out to assess changes in intravital biopsy specimens, is light microscopy, which in clinical practice makes it possible to establish the activity of reproduction.
burning and the stage of chronicity. The disadvantage of the method is the low resolution, which does not allow evaluating the structural features of cells, intracellular organelles, inclusions, and functional characteristics. Lifetime electron microscopic examination of ultrastructural changes in the liver makes it possible to supplement the data of light microscopy and increase their diagnostic value.
In this regard, the identification of hepatic HCI, the study of their phenotype in the process of transdifferentiation, and the determination of the intensity of their proliferation are the most important contribution to predicting the outcomes of liver diseases, as well as to the pathomorphology and pathophysiology of fibrogenesis.
Purpose - to present the structural and functional characteristics of HCI based on the results of cytological identification of intravital liver biopsy specimens.
Materials and methods
An intravital liver biopsy was obtained by aspiration liver biopsy in patients with CHC (HCV+ RNA), from whom written informed consent was obtained.
For light microscopy of semi-thin sections, liver biopsy samples of patients with a size of 0.5 × 2 mm were fixed by double fixation: first, according to the Sato Taizan method, then tissue samples were additionally fixed for 1 hour in a 1% osmium fixative prepared on 0.1 M phosphate Sorensen's buffer, pH 7.4. Potassium dichromate (K2Cr2O7) or chromic anhydride crystals (1 mg/mL) were added to 1% osmium tetroxide to better reveal intracellular structures and interstitial substance in semithin sections. After dehydration of the samples in a series of alcoholic solutions of increasing concentration and acetone, they were placed in a prepolymerized mixture of butyl methacrylate and styrene and polymerized at 550C. Semi-thin sections (1 µm thick) were sequentially stained
azure II-basic fuchsin. Micrographs were obtained using a digital video camera (Leica FC 320, Germany).
An electron microscopic study was carried out in samples of liver biopsy specimens 0.5x1.0 mm in size, fixed with a 1% solution of osmium tetroxide in 0.1 M Millonig's buffer, pH 7.4, at +40C for 2 hours. After dehydration in ascending alcohols and acetone, the samples were poured into araldite. Semithin sections (400 nm) were prepared from the obtained blocks on a Leica EM VC7 ultramicrotome (Germany) and stained with methylene blue. The preparations were examined under a light microscope and a single-type site was selected for further study of ultrastructural changes. Ultrathin sections (35 nm) were counterstained with 2% uranyl acetate in 50% methanol and lead citrate according to E. S. Reynolds. Electron microscopic preparations were studied using a JEM-1011 electron microscope (JEOL, Japan) at magnifications of 10,000–60,000 at an accelerating voltage of 80 kW. To obtain images, a complex from an Olympus MegaViewIII digital camera (Germany) and iTEM image processing software (Olympus, Germany) was used.
Results and discussion
HSCs are located in the perisinusoidal space (Disse) in pockets between hepatocytes and endothelial cells; they have long processes penetrating deep between hepatocytes. In most publications devoted to this population of HSCs, their schematic representation is given, which only allows one to designate the “territorial” belonging of HSCs in the liver and in relation to their surrounding “neighbors” (Figure 1).
HSCs are in close contact with endothelial cells through the components of an incomplete basement membrane and interstitial collagen fibers. Nerve endings penetrate between the SC and parenchymal cells, which is why the space of Disse is defined as the space between the plates of parenchymal cells and
a complex of HCI and endothelial cells.
HSCs are believed to originate from poorly differentiated mesenchymal cells in the transverse septum of the developing liver. The experiment found that hematopoietic stem cells are involved in the formation of HSCs and that this process is not due to cell fusion.
Sinusoidal cells (SCs), primarily HSCs, play a leading role in all types of liver regeneration. Fibrosing regeneration of the liver occurs as a result of inhibition of the stem functions of HSC and bone marrow stem cells. In the human liver, HSCs make up 5-15%, being one of the 4 varieties of SCs of mesenchymal origin: Kupffer cells, endotheliocytes, and Pb cells. The SC pool also contains 20-25% of leukocytes.
In the cytoplasm of HCI there are fatty inclusions with retinol, triglycerides, phospholipids, cholesterol, free fatty acids, a-actin and desmin. ZKI is visualized using gold chloride staining. It was found in the experiment that the marker of HKI differentiation from other myofibroblasts is their expression of the reelin protein.
HSCs exist in a quiescent ("inactive HSC"), transient and long-term activated state, each of which is characterized by gene expression and phenotype (α-IgMA, ICAM-1, chemokines and cytokines).
HSCs in an inactive state have a rounded, slightly elongated or irregular shape, a large nucleus and a bright visual sign - lipid inclusions (drops) containing retinol (Figure 2).
The number of lipid droplets in an inactive HSC reaches 30 or more, they are close in size, adjacent to each other, pressing into the nucleus and pushing it to the periphery (Figure 2). Small inclusions can be located between large drops. The color of the drops depends on the fixative and the color of the material. In one case, they are light (Figure 2a), in the other they are dark green (Figure 2b).
Figure 1. Scheme of the location of the ICH (stellatecell, perisinusoidal lipocyte) in the perisinusoidal space of Disse (space of Disse), Internet resource
Figure 2. - CCIs that are in an inactive state
a - round-shaped HCI with a high content of light-colored lipid droplets (white arrows), hepatocytes (Hz) with devastated cytoplasm (black arrow); b - HCI with dark lipid droplets in close contact with a macrophage (Mf); a-b - semi-thin sections. Coloring azure II - basic magenta. Micrographs. Increased 1000; c - HCI with an abundance of lipid droplets (more than 30), having an irregular shape (magnitude 6,000); d-ultrastructural components of HCI: l-lipid drops, mitochondria (orange arrows), GRES (green arrows), Golgi complex (red arrow), sw. 15,000; c-d - electronograms
With electron microscopy, a more osmiophilic marginal rim is formed against the background of a light lipid substrate (Figure 5a). In most "resting" HSCs, along with large lipid inclusions, there is a noticeably small amount of cytoplasmic matrix, poor in mitochondria (Mx) and granular endoplasmic reticulum (GRES). At the same time, compartments of a moderately developed Golgi complex are clearly visible in the form of a stack of 3-4 flattened cisterns with slightly widened ends (Figure 2d).
Under certain conditions, activated HSCs acquire a mixed or transitional phenotype, combining morphological features of both lipid-containing and fibroblast-like cells (Figure 3).
The transitional phenotype of HCI also has its own morphological features. The cell acquires an elongated shape, the number of lipid inclusions decreases, and the number of nucleolemma invaginations decreases. The volume of the cytoplasm increases, containing numerous GRES cisterns with bound ribosomes and free ribosomes, Mx. There is hyperplasia of the components of the lamellar Golgi complex, represented by several stacks of 3-8 flattened cisterns, an increase in the number of lysosomes involved in degradation
Figure 3. - ZKI, which are in a transitional state
a - ZKI (white arrows). Half cut. Coloring azure II - basic magenta. Micrograph. Increased 1000; b - ZKI of an elongated shape and with a small amount of lipid droplets; uv. 8000; c - HCI in contact with Kupffer cells (CC) and lymphocyte (Lc), SW. 6000. (Hz - hepatocyte, l - lipid drops, E - erythrocyte); d - mitochondria (orange arrows), GRES (green arrows), c. Goldji (red arrow), lysosomes (blue arrows), magn. b, c, d - electron diffraction patterns
lipid droplets (Figure 3d). Hyperplasia of the GRES components and the Golgi complex is associated with the ability of fibroblasts to synthesize collagen molecules, as well as to model them by post-translational hydroxylation and glycosylation in the endoplasmic reticulum and elements of the Golgi complex.
In an intact liver, HCI, being in a calm state, cover the sinusoidal capillary with their processes. The processes of HCI are divided into 2 types: perisinusoidal (subendothelial) and interhepatocellular (Figure 4).
The former leave the cell body and extend along the surface of the sinusoidal capillary, covering it with thin finger-like branches. They are covered with short villi and have characteristic long microprotrusions extending even further along the surface of the capillary endothelial tube. Interhepatocellular outgrowths, having overcome the plate of hepatocytes and reaching the neighboring sinusoid, are divided into several perisinusoidal outgrowths. Thus, the FQI covers on average more than two adjacent sinusoids.
With liver damage, the activation of HSC and the process of fibrogenesis occurs, in which 3 phases are distinguished. They are referred to as initiation, prolongation and resolution (resolution of fibrous tissue). This process of transformation of "resting" HSCs into fibrosing myofibroblasts is initiated by cytokines (^-1, ^-6,
Figure 4. - Perisinusoidal (subendothelial) and interhepatocellular processes (outgrowths) of HCI
(a) the process of the ZKI (yellow arrows) emerging from the cell body, uv. 30,000; b - a process of the HCI, located along the surface of the sinusoidal capillary, containing a lipid drop, SW. 30,000; (c) subendothelially located processes of the HCI. Processes of endothelial cells (pink arrows); d - interhepatocellular process of HCI; area of destruction of the membranes of the HCI and hepatocyte (black arrows), swollen 10 000. Electronograms
TOT-a), underoxidized metabolic products, reactive oxygen species, nitric oxide, endothelin, platelet activating factor (PDGF), plasminogen activator, transforming growth factor (TGF-1), acetaldehyde, and many others. Direct activators are hepatocytes in a state of oxidative stress, Kupffer cells, endotheliocytes, leukocytes, platelets producing cytokines (paracrine signals) and the ZKI itself (autocrine stimulation). Activation is accompanied by the expression (inclusion into work) of new genes, the synthesis of cytokines and proteins of the extracellular matrix (collagens I, III, Y types).
At this stage, the process of activation of HSCs can be completed by stimulating the formation of anti-inflammatory cytokines in HSCs, which inhibit the production of TOT-a by macrophages in the damaged area. As a result, the number of HSCs is sharply reduced, they undergo apoptosis, and fibrosis processes in the liver do not develop.
In the second phase (prolonged), with prolonged constant paracrine and autocrine exposure to activating stimuli, an activated phenotype is “maintained” in HSC, characterized by the transformation of HSC into contractile myofibroblast-like cells that synthesize extracellular fibrillar collagen.
The activated phenotype is characterized by proliferation, chemotaxis, contractility, loss of retinoid stores, and the formation of cells resembling myofibroblastic cells. Activated HSCs also show increased levels of new genes such as a-SMA, ICAM-1, chemokines and cytokines. Cell activation indicates the onset of an early stage of fibrogenesis and precedes increased production of ECM proteins. The resulting fibrous tissues undergo remodeling due to matrix cleavage with the help of matrix metalloproteinases (matrixmetaloproteinases - MMPs). In turn, matrix breakdown is regulated by tissue inhibitors of MMPs (tissue inhibitors of matrix metaloproteinases - TIMPs). MMPs and TIMPs are members of the zinc dependent enzyme family. MMPs are synthesized in HSCs as inactive proenzymes that are activated upon propeptide cleavage but are inhibited upon interaction with endogenous TIMPs, TIMPs-1 and TIMPs-2. HSCs produce 4 types of membrane-type MMPs that are activated by IL-1 p. Among MMPs, MMPs-9, a neutral matrix metalloproteinase, is of particular importance, which has activity against type 4 collagen, which is part of the basement membrane, as well as against partially denatured type 1 and 5 collagens.
An increase in the HCI population in various types of liver damage is judged by the activity of a significant number of mitogenic factors, related tyrosine kinase receptors and other identified mitogens that cause the most pronounced proliferation of HKI: endothelin-1, thrombin, FGF - fibroblast growth factor, PDGF - endothelial growth factor vessels, IGF - insulin-like growth factor. Accumulation of HSCs in areas of liver damage occurs not only due to the proliferation of these cells, but also due to their directed migration to these zones by chemotaxis, with the participation of chemoattractants such as PDGF and leukocyte chemoattractant-MCP (monocyte chemotactic protein-1) .
In activated HSCs, the number of lipid droplets is reduced to 1-3 with their location at opposite poles of the cell (Figure 5).
Activated HSCs acquire an elongated shape, significant areas of the cytoplasm are occupied by the Golgi complex, and quite numerous GRES cisterns (an indicator of protein synthesis for export) are revealed. The number of other organelles is reduced: few free ribosomes and polysomes, single mitochondria, and irregular lysosomes are found (Figure 6).
In 2007, HSCs were first named liver stem cells, since they express one of the markers of hematopoietic mesenchymal stem cells, CD133.
Figure 5. - CCIs in the activated state
a, b - HCI (blue arrows) with single lipid inclusions localized at opposite poles of the nucleus. The perisinusoidal connective tissue (Fig. 6a) and the intercellular matrix layer around the hepatocyte (Fig. 6b) are stained red. Cytotoxic lymphocytes (purple arrows). Endothelial cell (white arrow). Close contact between a plasma cell (red arrow) and a hepatocyte. Semi-thin cuts. Coloring azure II - basic magenta. Micrographs. Increased 1000 ; c, d - ultrastructural components of HCI: mitochondria (orange arrows), Golgi complex (red arrow), cisternae of its more osmiophilic cis-side facing extended elements of the granular endoplasmic reticulum (green arrows), lysosome (blue arrow) (magn. 10,000 and 20,000, respectively); c, d - electron diffraction patterns
Myofibroblasts, which are absent in the normal liver, have three potential sources: first, during intrauterine development of the liver, in the portal tracts, myofibroblasts surround the vessels and bile ducts during their maturation, and after full development of the liver, they disappear and are replaced in the portal tracts by portal fibroblasts; the second - with liver damage, they are formed due to portal mesenchymal cells and resting HSCs, less often due to transitional epithelial-mesenchymal cells. They are characterized by the presence of CD45-, CD34-, Desmin+, glial fibrillar protein associated with (GFAP)+ and Thy-1+.
Recent studies have shown that hepatocytes, cholangiocytes, and endothelial cells can become myofibroblasts via epithelial or endothelial-to-mesenchymal transition (EMT). These cells include markers such as CD45-, albumin+ (ie hepatocytes), CD45-, CK19+ (ie cholangiocytes) or Tie-2+ (endothelial cells).
Figure 6. - High fibrotic activity of HSC
a, b - myofibroblast (Mfb), the cell contains a large nucleus, GRES elements (red arrows), numerous free ribosomes, polymorphic vesicles and granules, single mitochondria and a bright visualization sign - a bundle of actin filaments in the cytoplasm (yellow arrows); led away. 12,000 and 40,000; c, d, e, f - high fibrotic activity of HSC with retention of retinoid-containing lipid droplets in the cytoplasm. Numerous bundles of collagen fibrils (white arrows) retained (a) and lost (d, e, f) specific transverse striation; led away. 25,000, 15,000, 8,000, 15,000. Electronograms
In addition, bone marrow cells, consisting of fibrocytes and circulating mesenchymal cells, can transform into myofibroblasts. These are CD45+ (fibrocytes), CD45+/- (circulating mesenchymal cells), collagen type 1+, CD11d+ and MHC class 11+ (Figure 7).
Literature data confirm not only the close relationship between the proliferation of oval cells and the proliferation of sinusoidal cells, but also data on the possible differentiation of HSC into the hepatic epithelium, which was called mesenchymal-epithelial transformation of perisinusoidal cells.
In the state of fibrogenic activation, myofibroblast-like HSCs, along with a decrease in the number and subsequent disappearance of lipid droplets, are characterized by focal proliferation (Figure 8), immunohistochemical expression of fibroblast-like markers, including smooth muscle α-actin, and the formation of pericellular collagen fibrils in the spaces of Disse.
In the phase of fibrosis development, the increasing hypoxia of the liver tissue becomes a factor of additional overexpression in stem cells of pro-inflammatory adhesion molecules - 1CAM-1, 1CAM-2, VEGF, pro-inflammatory
Interaction of ductal hepatic progenitor cells with liver myofibroblasts
Myofibroblast-like HSCs in a state of fibrogenic activation.
Figure 7. - Participants of myofibroblastic activation of HSC
potent chemoattractants - M-CSF, MCP-1 (monocyte chemotactic protein-1) and SGS (cytokine-mediated neutrophil chemoattractant) and others that stimulate the formation of pro-inflammatory cytokines (TGF-b, PDGF, FGF, PAF, SCF, ET-1 ) and enhance the processes of fibrogenesis in the liver, creating conditions for self-sustained induction of the ongoing activation of HSC and fibrogenesis processes.
On microscopic preparations, pericapillary fibrosis manifests itself in the form of intense coloring of the perisinusoidal connective tissue and the intercellular matrix layer around hepatocytes (often dying) in red. On electron microscopic preparations, fibrotic changes are visualized either in the form of formed large bundles of fibrils of collagen fibers that have retained transverse striation, or in the form of a massive
deposits in the space of Disse fibrous mass, which is swollen collagen fibers that have lost their periodic striation (Figure 9).
According to modern concepts, fibrosis is a dynamic process that can progress and regress (Figure 10).
Recently, several specific markers of ICD have been proposed: vitamin A (VA) bloom in lipid droplets, GFAP, the p75 NGF receptor, and synaptophysin. Studies are being conducted on the involvement of liver HCI in the proliferation and differentiation of liver stem cells.
We have studied the content of retinol-binding protein (RBP-4), which forms a complex with VA, the concentration of which in the blood plasma normally correlates with the provision of the body with VA, 80% of which is in the HCI.
The relationship between the content
Figure 8. - Focal proliferation of HSC in the state of fibrogenic activation
a - HCI hyperplasia (white arrows) in the lumen of dilated sinusoids; b - proliferation of transdifferentiated HSCs (white arrows), endothelial cell (pink arrow). Semi-thin cuts. Coloring azure II - basic magenta. Micrographs. Increased 1000
Figure 9. - The final stage of myofibroblastic activation of HSC
a, b - perisinusoidal fibrosis (white arrows). The peri-sinusoidal connective tissue and the intercellular matrix layer around the hepatocytes (b) are stained red with basic fuchsin. HSCs activated and transformed into fibroblasts (blue arrows). Hz in fig. a - hepatocyte with devastated cytoplasm. Semi-thin cuts. Coloring azure II - basic magenta. Micrographs. Increased 1000; c, d - perisinusoidal and perihepatocellular fibrosis in the liver lobule, increased electron density of collagen fiber fibrils; condensation of the mitochondrial matrix in the hepatocyte (orange arrow). Boost 8,000 and 15,000, respectively. electronograms
Table 1. Indicators of RBP-4 content in patients with liver cirrhosis (LC) and chronic hepatitis (CH) of various etiologies, ng/ml (M±m)
Group n M±m p
Liver cirrhosis 17 23.6±2.29<0,05
CG, AsAT norm 16 36.9±2.05* >0.05
CG, ASAT >2 norms 13 33.0±3.04* >0.05
CG, ALT norm 13 37.5±3.02* >0.05
CG, ALT >2 norms 21 35.9±2.25* >0.05
Control 15 31.2±2.82
Note: p - significant differences with control (p<0,05); * - достоверные различия между ЦП и ХГ (р<0,05)
False lobule surrounded by a fibrous septum with a fibrous septum. Coloring according to Masso - a circle of a false lobule. Coloring according to u.Uv.x50 Masson. Increase x200
Figure 10 - Dynamics of events in the false lobule of a patient with viral cirrhosis 6 months after transplantation of autologous mesenchymal stem cells into the liver
I eat RBP-4 and stage 4 fibrosis (cirrhosis), in contrast to chronic hepatitis, in which such a dependence was not observed, regardless of biochemical markers of inflammation activity in the liver.
This fact must be taken into account when substantiating replacement therapy to eliminate VA deficiency in the body, which may be due to the depletion of the potential of HSC due to the progression of fibrosis in the liver.
1. The maximum effectiveness of the assessment of the structural and functional state of HCI is ensured by a morphological study of an intravital biopsy specimen with the simultaneous use of a complex of cell visualization techniques (light, electron microscopy of ultrathin sections and original methods of fixation and staining).
2. The results of the morphological study of HCI allow to improve the quality of in vivo diagnosis of fibrosis, to monitor it and predict the outcomes of chronic diffuse liver lesions at a higher modern level.
3. The results of morphological conclusions will allow the clinician to additionally include refined data on the stage of chronicity (stabilization, progression or resolution of fibrosis) in the course of therapy in the formulation of the final diagnosis.
Literature
1. Ivashkin, V. T. Clinical symptoms of pre-fibrotic changes: a transcript of the lecture of the All-Russian Internet Congress of Internal Medicine Specialists / V. T. Ivashkin, A. O. Bueverov // INTERNIST: National Internet Society of Internal Medicine Specialists. - 2013. - Access mode: http://internist. ru/publications/detail/6569/. - Date of access: 21.11.2016.
2. Kiyasov, A.P. Oval cells - putative liver stem cells or hepatoblasts? / A. P. Kiyasov, A. A. Gumerova, M. A. Titova // Cell transplantology and tissue engineering. - 2006. - V. 2, No. 4. - S. 55-58.
1. Ivashkin, V. T. Klinicheskaya simptomatika dofibroticheskih izmenenij: stenogramma lekcii Vserossijskogo Internet-Kongressa specialistov po vnutrennim boleznyam / V. T. Ivashkin, A. O. Bueverov // INTERNIST: Nacional "noe Internet-Obshchestvo specialistov po vnutrennim boleznyam. - http: Rezhipa 20 do13. - http: Rezhipa 20 do13. - //internist.ru/publications/detail/6569/ - Data accessed: 11/21/2016.
2. Kiyasov, A. P. Oval "nye kletki - predpolagaemye stvolovye kletki pecheni or gepatoblasty? / A. P. Kiyasov, A. A. Gumerova, M. A. Titova // Kletochnaya transplantologiya i tkanevaya inzheneriya. - 2006. - T. 2, No. 4. - S. 55- 58.
3. On the role of sinusoidal liver cells and bone marrow cells in providing a regenerative strategy for a healthy and damaged liver / A. V. Lundup [et al.] // Bulletin of transplantology and artificial organs. -2010. - T. XII, No. 1. - S. 78-85.
4. Serov, V. V. Morphological criteria for assessing the etiology, degree of activity and stage of the process in viral chronic hepatitis B and C / V. V. Serov, L. O. Severgina // Archives of Pathology. - 1996. - No. 4. - S. 61-64.
5. Structural and functional characteristics of liver stellate cells in the dynamics of fibrosis / OA Postnikova [et al.] // Fundamental research. - 2011. - No. 10.
6. Ultrastructural and immunohistochemical study of liver stellate cells in the dynamics of fibrosis and cirrhosis of the infectious-viral genesis / G. I. Nepomnyashchikh [et al.] // Bulletin of experimental biology and medicine. - 2006. - T. 142, No. 12. - S. 681-686.
7. Shcheglev, AI Structural and metabolic characteristics of sinusoidal liver cells / AI Shcheglev, OD Mishnev // Successes of modern biology. - 1991. - V. 3, No. 1. - S. 73-82.
10. Effects of dietary retinoid and triglyceride on the lipid composition of rat liver stellate cells and stellate cell lipid droplets / H. Moriwaki // J. Lipid. Res. - 1988. - Vol. 29. - R. 1523-1534.
13. Friedman, S. Hepatic fibrosis 2006: Report of the Third AASLD Single Topic Conference / S. Friedman, D. Rockey, B. Montgomery // Hepatology. - 2006. - Vol. 45(1). - R. 242-249.
18. Iredale, J. P. Hepatic Stellate Cell Behavior During Resolution of Liver Injury / J. P. Iredale // Semin. LiverDis. -2001. - Vol. 21(3). - R. 427-436.
19. Kobold, D. Expression of reelin in hepatic stellate cells and during hepatic tissue repair: a novel marker for the differentiation of HSC from other liver myofibroblasts / D. Kobold // J. Hepatol. - 2002. - Vol. 36(5). - R. 607-613.
20. Lepreux, S. Human liver myofibroblasts during development and diseases with a focus on portal (myo)
3. O roli sinusoidal "nyh kletok pecheni i kletok kostnogo mozga v obespechenii regeneratornoj strategii zdorovoj i povrezhdennoj pecheni / A. V. Lyundup // Vestnik transplantologii i iskusstvennyh organov. - 2010. - T. HII, No. 1. - S. 78-85.
4. Serov, V. V. Morfologicheskie kriterii ocenki ehtiologii, stepeni aktivnosti i stadii processa pri virusnyh chrononicheskih gepatitah V i S / V. V. Serov, L. O. Severgina // Arhiv patologii.
1996. - No. 4. - S. 61-64.
5. Strukturno-funkcional "naya harakteristika zvezdchatyh kletok pecheni v dinamike fibroza / O. A. Postnikova // Fundamental" nye issledovaniya. - 2011. - No. 10. - C. 359-362.
6. Ul "trastrukturnoe i immunogistohimicheskoe issledovanie zvezdchatyh kletok pecheni v dinamike fibroza i cirroza pecheni infekcionno-virusnogo geneza / G. I. Nepomnyashchih // Byulleten" ehksperimental "nojologii i mediciny. - 2006. - T. 142, No. 681. - S. -686.
7. SHCHeglev, A. I. Strukturno-metabolicheskaya harakteristika sinusoidal "nyh kletok pecheni / A. I. SHCHeglev, O. D. Mishnev // Uspekhi sovremennoj biologii. - 1991. - T. 3, No. 1. - S. 73-82.
8. CD34 hepatic stellate cells are progenitor cells / C. Kordes // Biochem., Biophys. Res. Common. - 2007. -Vol. 352(2). - P. 410-417.
9. Degradation of matrix proteins in liver fibrosis / M. J. Arthur // Pathol. Res. Pract. - 1994. - Vol. 190(9-10).
10. Effects of dietary retinoid and triglyceride on the lipid composition of rat liver stellate cells and stellate cell lipid droplets / H. Moriwaki // J. Lipid. Res. - 1988. - Vol. 29. - R. 1523-1534.
11. Fetal liver consists of cells in epithelial-to-mesenchymal transition / J. Chagraoni // Blood. - 2003. - Vol. 101. - P. 2973-2982.
12. Fixation, dehydration and embedding of biological specimens / A. M. Glauert // Practical Methods in Electron Microscopy. - New York: Am. Elsevier, 1975. - Vol. 3, part 1.
13. Friedman, S. Hepatic fibrosis 2006: Report of the Third AASLD Single Topic Conference / S. Friedman, D. Rockey, B. Montgomery // Hepatology. - 2006. - Vol. 45(1). - R. 242-249.
14. Gaga, M. D. Human and rathepatic stellate cells produce stem cell factor: a possible mechanism for mast cell recruitment in liver fibrosis / M. D. Gaga // J. Hepatol. - 1999. - Vol. 30, No. 5. - P. 850-858.
15. Glauert, A. M. Araldite as embedding medium for electron microscopy / A. M. Glauert, R. H. Glauert // J. Biophys. Biochem. Cytol. - 1958. - Vol. 4. - P. 409-414.
16. Hepatic stellate cells and portal fibroblasts are the major cellular sources of collagens and lysyl oxidases in normal liver and early after injury / M. Perepelyuk // Am. J Physiol. gastrointest. Liver Physiol. - 2013. - Vol. 304(6). - P. 605614.
17. Hepatitis C virus core and nonstructural proteins induce fibrogenic effects in hepatic stellate cells / R. Bataller // Gastroenterology. - 2004. - Vol. 126, iss. 2. - P. 529-540.
18. Iredale, J. P. Hepatic Stellate Cell Behavior During Resolution of Liver Injury / J. P. Iredale // Semin. LiverDis. -2001. - Vol. 21(3). - R. 427-436.
19. Kobold, D. Expression of reelin in hepatic stellate cells and during hepatic tissue repair: a novel marker for the differentiation of HSC from other liver myofibroblasts / D. Kobold // J. Hepatol. - 2002. - Vol. 36(5). - R. 607-613.
20. Lepreux, S. Human liver myofibroblasts during development and diseases with a focus on portal (myo) fibroblasts / S. Lepreux, A. Desmouliére
fibroblasts / S. Lepreux, A. Desmoulière // Front. physiol. - 2015. - Mode of access: http://dx.doi. org/10.3389/fphys.2015.00173. - Date of access: 31.10.2016.
22. Mesenchymal Bone Marrow-derived Stem Cells Transplantation in Patients with HCV Related Liver Cirrhosis / S. Lukashyk // J. Clin. Transl. Hepatol. - 2014. - Vol. 2, iss. 4. - P. 217-221.
23. Millonig, G. A. Advantages of a phosphate buffer for osmium tetroxide solutions in fixation / G. A. Millonig // J. Appl. Physics. - 1961. - Vol. 32. - P. 1637-1643.
Vol. 158. - P. 1313-1323.
Vol. 24. - P. 205-224.
29. Querner, F. Der mikroskopische Nachweis von Vitamin Aimanimalen Gewebe. Zur Kenntnis der paraplasmatischen Leberzellen-einschlüsse. Dritte Mitteilung / F. Querner // Klin. Wschr. - 1935. - Vol. 14. - P. 1213-1217.
30. Recent developments in myofibroblast biology: paradigms for connective tissue remodeling / B. Hinz // Am. J. Pathol. - 2012. - Vol. 180. - P. 1340-1355.
35. Septum transversum-derived mesothelium gives rise to hepatic stellate cells and perivascular mesenchymal cells in developing mouse liver / K. Asahina // Hepatology. -2011. - Vol. 53.-P. 983-995.
Vol. 50.-P. 66-71.
38. Thabut, D. Intrahepatic angiogenesis and sinusoidal remodeling in chronic liver disease: new targets for the treatment of portal hypertension? / D. Thabut, V. Shah // J. Hepatol. - 2010. - Vol. 53. - P. 976-980.
39. Wake, K. Hepatic stellate cells: Three-dimensional structure, localization, heterogeneity and development / K.
// front. physiol. - 2015. - Mode of access: http://dx.doi. org/10.3389/fphys.2015.00173. - Date of access: 31.10.2016.
21. Ligands of peroxisome proliferator-activated receptor gamma mod-ulateprofibrogenic and proinflammatory actions in hepatic stellate cells / F. Marra // Gastroenterology. -2000. - Vol. 119. - P. 466-478.
22. Mesenchymal Bone Marrow-derived Stem Cells Transplantation in Patients with HCV Related Liver Cirrhosis / S. Lukashyk // J. Clin. Transl. Hepatol. - 2014. - Vol. 2, iss. 4.-R. 217-221.
23. Millonig, G. A. Advantages of a phosphate buffer for osmium tetroxide solutions in fixation / G. A. Millonig // J. Appl. Rhysics. - 1961. - Vol. 32. - P. 1637-1643.
24. Origin and structural evolution of the early proliferating oval cells in rat liver / S. Paku // Am. J. Hepatol. - 2001.
Vol. 158. - P. 1313-1323.
25. Origin of myofibroblasts in liver fibrosis / D. A. Brenner // Fibrogenesis Tissue Repair. - 2012. - Vol. 5 suppl. 1. - S. 17.
26. Origins and functions of liver myofibroblasts / S. Lemoinne // Biochim. Biophys. acta. - 2013. - Vol. 1832(7). - P. 948-954.
27. Pinzani, M. PDGF and signal transduction in hepatic stellate cells / M. Pinzani // Front. biosci. - 2002. - Vol. 7. - P. 1720-1726.
28. Popper, H. Distribution of vitamin A in tissue as revealed by fluorescence microscopy / H. Popper // Physiol. Rev. - 1944.
Vol. 24.-R. 205-224.
29. Querner, F. Der mikroskopische Nachweis von Vitamin Aimanimalen Gewebe. Zur Kenntnis der paraplasmatischen Leberzellen-einschlüsse. Dritte Mitteilung / F. Querner // Klin. Wschr. - 1935. - Vol. 14. - R. 1213-1217.
30. Recent developments in myofibroblast biology: paradigms for connective tissue remodeling / B. Hinz // Am. J. Pathol. - 2012. - Vol. 180. - R. 1340-1355.
31. Reynolds, E. S. The use of lead citrate at high pH as an electronopaque stain in electron microscopy / E. S. Reynolds // J. Cell. Biol. - 1963. - Vol. 17. - P. 208-212.
32. Safadi, R. Immune stimulation of hepatic fibrogenesis by CD8 cells and attenuation by transgenic interleukin-10 from hepatocytes / R. Safadi // Gastroenterology. - 2004. - Vol. 127(3). - P. 870-882.
33. Sato, T. An electron microscopic study of specimen-fixed for longer periods in phosphate buffered formalin / T. Sato, I. Takagi // J. Electron Microsc. - 1982. - Vol. 31, No. 4. - P. 423-428.
34. Senoo, H. Vitamin A-Storing Cells (Stellate Cells) / H. Senoo, N. Kojima, M. Sato // Vitam. Horm. - 2007. - Vol. 75.
35. Septum transversum-derived mesothelium gives rise to hepatic stellate cells and perivascular mesenchymal cells in developing mouse liver / K. Asahina // Hepatology. -2011. - Vol. 53.-R. 983-995.
36. Stanciu, A. New data about ITO cells / A. Stanciu, C. Cotutiu, C. Amalinei, Rev. Med. Chir. soc. Med. Nat. Iasi. -2002. - Vol. 107, No. 2. - P. 235-239.
37. Suematsu, M. Professor Toshio Ito: a clairvoyant in pericyte biology / M. Suematsu, S. Aiso // Keio J. Med. - 2000.
Vol. 50.-R. 66-71.
38. Thabut, D. Intrahepatic angiogenesis and sinusoidal remodeling in chronic liver disease: new targets for the treatment of portal hypertension? / D. Thabut, V. Shah // J. Hepatol. - 2010. - Vol. 53.-R. 976-980.
39. Wake, K. Hepatic stellate cells: Three-dimensional structure, localization, heterogeneity and development / K. Wake // Proc. Jpn. Acad. Ser. B, Phys. Biol. sci. - 2006. - Vol.
Wake // Proc. Jpn. Acad. Ser. B, Phys. Biol. sci. - 2006. - Vol. 82(4). - P. 155-164.
82(4). - P. 155-164.
40. Wake, K. In Cells of the Hepatic Sinusoid / K. Wake, H. Senoo // Kupffer Cell Foundation (Rijswijk, The Netherlands). - 1986. - Vol. 1. - P. 215-220.
41. Watson, M. L. Staining of tissue sections for electron micr with heavy metals / M. L. Watson // J. Biophys. Biochem. Cyt. - 1958. - Vol. 4. - P. 475-478.
CLINICAL CYTOLOGY OF THE LIVER: ITO STELLATE CELLS (HEPATIC STELLATE CELLS)
Tsyrkunov V. M, Andreev V. P., Kravchuk R. I., Kandratovich I. A. Educational Establishment "Grodno State Medical University", Grodno, Belarus
introduction. The role of Ito stellate cells (Hepatic Stellate Cells, HSC) has been identified as one of the leading in the development of liver fibrosis, but the use of intravital visualization of HSC structures in clinical practice is minimal.
The aim of the work is to present the structural and functional characteristic of HSC based on the findings of cytological identification of intravital liver biopsy samples.
materials and methods. Classical methods of light and electron microscopy of biopsy samples within the original technique of using ultrathin sections, fixation and staining were applied.
results. The structural characteristics of the HSC of the liver biopsy samples from patients with chronic hepatitis C are presented on photo illustrations of light and electron microscopy. HSCs are depicted at different stages (rest, activation) and during the process of transformation into myofibroblasts.
Conclusions. The use of original methods of clinical and morphological identification and evaluation of the functional status of HSC allows to improve the quality of diagnosis and prognosis of liver fibrosis.
In this case, these cells respond by proliferating to the effects of cytokines, growth factors, and chemokines (pro-inflammatory cytokines) produced by the damaged liver. Chronic activation of stellate cells in response to oxidative stress caused by HBV and HCV replication may contribute to fibrogenesis and increased proliferation of hepatocytes chronically infected with HBV and HCV.
Thus, stellate cells are involved in the regulation of growth, differentiation, and circulation of hepatocytes, which, together with the activation of MAP kinases, can lead to the development of liver cancer [Block, 2003].
Links:
Random drawing
Attention! Information on the site
intended solely for educational
Studying the effect of liver Ito cells on stem cells
Intercellular communication might be realized by paracrine secretion and direct cell-to-cell contacts. It is known that hepatic perisinusoidal cells (HPC) establish regional stem cells niche and determine their differentiation. At the same time HPC remain poorly characterized on the molecular and cellular level.
Shafigullina A.K., Trondin A.A., Shaikhutdinova A.R., Kaligin M.S., Gazizov I.M., Rizvanov A.A., Gumerova A.A., Kiyasov A.P.
SEI HPE "Kazan State Medical University of the Federal Agency for Health and Social Development"
Experimental Evaluation of the Osteoinductance of a Recombinant Bone Morphogenetic Protein
Cell technologies in the treatment of degenerative-dystrophic diseases of bones and joints
Ito cage
calm and activated. Activated Ito cells
calm state
perisinusoidal(subendothelial) and interhepatocellular. The former leave the cell body and extend along the surface of the sinusoidal capillary, covering it with thin finger-like branches. Perisinusoidal outgrowths are covered with short villi and have characteristic long microprotrusions extending even further along the surface of the capillary endothelial tube. Interhepatocellular outgrowths, having overcome the plate of hepatocytes and reaching the neighboring sinusoid, are divided into several perisinusoidal outgrowths. Thus, the Ito cell covers, on average, slightly more than two adjacent sinusoids.
activated state
liver cells
The human liver consists of cells, like any organic tissue. Nature is arranged in such a way that this organ performs the most important functions, it cleanses the body, produces bile, accumulates and deposits glycogen, synthesizes plasma proteins, manages metabolic processes, participates in the normalization of the amount of cholesterol and other components necessary for the life of the body.
To fulfill their purpose, liver cells must be healthy, have a stable structure, each person needs to protect them from destruction.
On the structure and types of hepatic lobules
The cellular composition of the body is characterized by diversity. Liver cells make up lobules, segments are made up of lobules. The structure of the organ is such that hepatocytes (the main liver cells) are located around the central vein, branch off from it, connect with each other, forming sinusoids, that is, gaps filled with blood. Blood moves through them like capillaries. The liver is supplied with blood from the portal vein and artery located in the organ. The liver lobules produce bile and carry it into the bile ducts.
Other types of liver cells and their purpose
- Endothelial - cells lining the sinusoids and containing fenestra. The latter are designed to form a stepped barrier between the sinusoid and the Disse space.
- The Disse space itself is filled with stellate cells; they ensure the outflow of tissue fluid into the lymphatic vessels of the portal zones.
- Kupffer cells are associated with the endothelium, they are attached to it, their function is to protect the liver when a generalized infection enters the body, in case of injury.
- Pit cells are killers of hepatocytes affected by the virus, in addition, they have cytotoxicity to tumor cells.
The human liver consists of 60% hepatocytes and 40% other types of cellular compounds. Hepatocytes look like a polyhedron, there are at least 250 billion of them. The normal functioning of hepatocytes is due to the spectrum of components that are secreted by the sinusoidal cells that fill the sinusoidal compartment. That is, the above Kupffer, stellate and pit cells (intrahepatic lymphocytes).
Endothelial cells are a filter between blood in the sinusoidal space and plasma in the Disse space. This biological filter sorts out large, excessively rich in retinol and cholesterol compounds and does not let them through, which is beneficial for the body. In addition, their function is to protect the liver (namely, hepatocytes) from mechanical damage by blood cells.
Our regular reader recommended an effective method! New discovery! Novosibirsk scientists have identified the best remedy for cleansing the liver. 5 years of research. Self-treatment at home! After carefully reviewing it, we decided to offer it to your attention.
The process of interaction of the elements of the body
Between all the particles of the body there is an interaction that has a rather complex scheme. A healthy liver is characterized by the stability of cellular compounds; in pathological processes, an extracellular matrix can be traced under a microscope.
Organ tissue under the influence of toxins, such as alcohol, viral agents, undergoes changes. They are as follows:
- deposition in the body of products resulting from metabolic disorders;
- cell dystrophy;
- necrosis of hepatocytes;
- fibrosis of hepatic tissues;
- inflammatory process of the liver;
- cholestasis.
About the treatment of organ pathology
It is useful for every patient to know what the changes that an organ undergoes mean. Not all of them are catastrophic. For example, dystrophy can be mild or severe. Both of these processes are reversible. Currently, there are drugs that restore cells and entire segments of the liver.
Cholestasis can be cured even with folk remedies - decoctions and infusions. They contribute to the normalization of the synthesis of bilirubin and eliminate disturbances in the outflow of bile into the duodenum.
With cirrhosis in the initial stage, treatment begins with a diet, then therapy with hepatoprotectors is prescribed. The most effective way to treat cirrhosis and fibrosis are stem cells, which are injected into the umbilical vein or intravenously, they restore hepatocytes damaged by various agents.
The main causes of liver cell death are alcohol abuse, drug exposure, including drugs, medicines. Any toxin that enters the body is a liver destroyer. Therefore, you should give up bad habits so that you have a healthy liver.
Who said that it is impossible to cure severe liver diseases?
- Many methods have been tried, but nothing helps.
- And now you are ready to take advantage of any opportunity that will give you the long-awaited good health!
An effective remedy for the treatment of the liver exists. Follow the link and find out what the doctors recommend!
Read also:
Education: Rostov State Medical University (RostGMU), Department of Gastroenterology and Endoscopy.
ENDOTHELIAL CELLS, KUPFER AND ITO CELLS
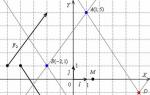
The structure of endothelial cells, Kupffer and Ito cells, we will consider on the example of two figures.
The figure to the right of the text shows the sinusoidal capillaries (SC) of the liver - intralobular capillaries of the sinusoidal type, increasing from the input venules to the central vein. The hepatic sinusoid capillaries form an anastomotic network between the hepatic laminae. The lining of sinusoidal capillaries is formed by endothelial cells and Kupffer cells.
In the figure to the left of the text, the liver lamina (LP) and two sinusoidal capillaries (SCs) of the liver are cut vertically and horizontally to show perisinusoidal Ito cells (CIs). The figure also shows cut bile ducts (LC).
ENDOTHELIAL CELLS
Endothelial cells (ECs) are highly flattened squamous cells with an elongated small nucleus, underdeveloped organelles, and a large number of micropinocytic vesicles. The cytomembrane is dotted with non-permanent holes (O) and fenestra, often grouped into cribriform plates (RP). These openings allow blood plasma to pass through, but not blood cells, allowing it access to hepatocytes (D). Endothelial cells do not have a basement membrane and do not possess phagocytosis. They are connected to each other using small connector complexes (not shown). Together with Kupffer cells, endothelial cells form the inner border of the space of Disse (PD); its outer border is formed by hepatocytes.
KUPFER CELLS
Kupffer cells (CC) are large, nonpermanent stellate cells within the hepatic sinusoidal capillaries, partly at their bifurcations.
The processes of Kupffer cells pass without any connecting devices between endothelial cells and often cross the lumen of the sinusoids. Kupffer cells contain an oval nucleus, many mitochondria, a well-developed Golgi complex, short cisterns of the granular endoplasmic reticulum, many lysosomes (L), residual bodies, and rare annular plates. Kupffer cells also contain large phagolysosomes (PL), which often contain obsolete erythrocytes and foreign matter. Hemosiderin or iron inclusions may also be detected, especially on supravital staining.
The surface of Kupffer cells shows irregular flattened cytoplasmic folds called lamellipodia (LP) - lamellar stalks, as well as processes called filopodia (F) and microvilli (MV) covered with glycocalyx. The plasmalemma forms vermiform bodies (CT) with a centrally located dense line. These structures may represent a condensed glycocalyx.
Kupffer cells are macrophages, most likely forming an independent cell genus. They usually originate from other Kupffer cells due to mitotic division of the latter, but may also originate from the bone marrow. Some authors believe that they are activated endothelial cells.
Occasionally, a random autonomic nerve fiber (NF) passes through the space of Disse. In some cases, the fibers have contact with hepatocytes. The edges of hepatocytes are delimited by interhepatocyte depressions (MU) dotted with microvilli.
ITO CELLS
These are stellate cells localized within the spaces of Disse (PD). Their nuclei are rich in condensed chromatin and are usually deformed by large lipid drops (LA). The latter are present not only in the perikaryon, but also in the processes of the cell and are visible from the outside as spherical protrusions. Organelles are poorly developed. Perisinusoidal cells show weak endocytic activity, but lack phagosomes. Cells have several long processes (O) that are in contact with neighboring hepatocytes, but do not form connecting complexes.
The processes enclose the sinusoidal capillaries of the liver and in some cases pass through the hepatic laminae, coming into contact with adjacent hepatic sinusoids. The processes are not constant, branched and thin; they may also be flattened. Accumulating groups of lipid drops, they lengthen and take on the appearance of a grape brush.
It is believed that perisinusoidal Ito cells are poorly differentiated mesenchymal cells that can be considered as hematopoietic stem cells, since they can transform under pathological conditions into fat cells, active blood stem cells, or fibroblasts.
Under normal conditions, Ito cells are involved in the accumulation of fat and vitamin A as well as in the production of intralobular reticular and collagen fibers (KB).
Psychology and psychotherapy
This section will include articles on research methods, drugs and other components related to medical topics.
A small section of the site that contains articles about original items. Watches, furniture, decorative items - all this you can find in this section. The section is not the main one for the site, and rather serves as an interesting addition to the world of human anatomy and physiology.
Ito liver cells
Universal Popular Science Online Encyclopedia
LIVER
LIVER, the largest gland in the body of vertebrates. In humans, it is about 2.5% of body weight, an average of 1.5 kg in adult men and 1.2 kg in women. The liver is located in the right upper part of the abdominal cavity; it is attached by ligaments to the diaphragm, abdominal wall, stomach and intestines and is covered with a thin fibrous membrane - the glisson capsule. The liver is a soft but dense organ of red-brown color and usually consists of four lobes: a large right lobe, a smaller left one, and much smaller caudate and square lobes that form the posterior lower surface of the liver.
Functions.
The liver is an essential organ for life with many different functions. One of the main ones is the formation and secretion of bile, a clear orange or yellow liquid. Bile contains acids, salts, phospholipids (fats containing a phosphate group), cholesterol, and pigments. Bile salts and free bile acids emulsify fats (i.e., break them into small droplets), which makes them easier to digest; convert fatty acids into water-soluble forms (which is necessary for the absorption of both the fatty acids themselves and fat-soluble vitamins A, D, E and K); have antibacterial action.
All the nutrients absorbed into the blood from the digestive tract - the products of digestion of carbohydrates, proteins and fats, minerals and vitamins - pass through the liver and are processed in it. At the same time, part of the amino acids (protein fragments) and part of the fats are converted into carbohydrates, so the liver is the largest "depot" of glycogen in the body. It synthesizes blood plasma proteins - globulins and albumin, as well as amino acid conversion reactions (deamination and transamination). Deamination - the removal of nitrogen-containing amino groups from amino acids - allows the latter to be used, for example, for the synthesis of carbohydrates and fats. Transamination is the transfer of an amino group from an amino acid to a keto acid to form another amino acid ( cm. METABOLISM). The liver also synthesizes ketone bodies (products of fatty acid metabolism) and cholesterol.
The liver is involved in the regulation of glucose (sugar) levels in the blood. If this level rises, liver cells convert glucose into glycogen (a substance similar to starch) and store it. If the blood glucose falls below normal, glycogen is broken down and glucose enters the bloodstream. In addition, the liver is able to synthesize glucose from other substances, such as amino acids; this process is called gluconeogenesis.
Another function of the liver is detoxification. Drugs and other potentially toxic compounds can be converted into a water-soluble form in the liver cells, which allows them to be excreted in the bile; they can also be destroyed or conjugated (combined) with other substances to form harmless products that are easily excreted from the body. Some substances are temporarily deposited in Kupffer cells (special cells that absorb foreign particles) or in other liver cells. Kupffer cells are particularly effective at removing and destroying bacteria and other foreign particles. Thanks to them, the liver plays an important role in the immune defense of the body. Possessing a dense network of blood vessels, the liver also serves as a reservoir of blood (it constantly contains about 0.5 liters of blood) and is involved in the regulation of blood volume and blood flow in the body.
In general, the liver performs more than 500 different functions, and its activity cannot yet be reproduced artificially. Removal of this organ inevitably leads to death within 1–5 days. However, the liver has a huge internal reserve, it has an amazing ability to recover from damage, so humans and other mammals can survive even after the removal of 70% of liver tissue.
Structure.
The complex structure of the liver is perfectly adapted to its unique functions. Shares consist of small structural units - lobules. In the human liver, there are about one hundred thousand of them, each 1.5–2 mm long and 1–1.2 mm wide. The lobule consists of liver cells - hepatocytes, located around the central vein. Hepatocytes unite in layers one cell thick - the so-called. liver plates. They diverge radially from the central vein, branch and connect with each other, forming a complex system of walls; narrow gaps between them, filled with blood, are known as sinusoids. Sinusoids are equivalent to capillaries; passing one into another, they form a continuous labyrinth. The hepatic lobules are supplied with blood from the branches of the portal vein and the hepatic artery, and the bile formed in the lobules enters the tubule system, from them into the bile ducts and is excreted from the liver.
The hepatic portal vein and hepatic artery provide the liver with an unusual, dual blood supply. Nutrient-rich blood from the capillaries of the stomach, intestines, and several other organs is collected in the portal vein, which, instead of carrying blood to the heart like most other veins, carries it to the liver. In the lobules of the liver, the portal vein breaks up into a network of capillaries (sinusoids). The term "portal vein" indicates an unusual direction of blood transport from the capillaries of one organ to the capillaries of another (the kidneys and pituitary gland have a similar circulatory system).
The second blood supply to the liver, the hepatic artery, carries oxygenated blood from the heart to the outer surfaces of the lobules. The portal vein provides 75-80%, and the hepatic artery 20-25% of the total blood supply to the liver. In general, about 1500 ml of blood passes through the liver per minute, i.e. quarter of cardiac output. Blood from both sources ends up in the sinusoids, where it mixes and goes to the central vein. From the central vein begins the outflow of blood to the heart through the lobar veins into the hepatic (not to be confused with the portal vein of the liver).
Bile is secreted by the liver cells into the smallest tubules between cells - the bile capillaries. Through the internal system of tubules and ducts, it is collected in the bile duct. Some of the bile goes straight into the common bile duct and out into the small intestine, but most of it is returned through the cystic duct to the gallbladder, a small, muscular sac attached to the liver, for storage. When food enters the intestine, the gallbladder contracts and ejects the contents into the common bile duct, which opens into the duodenum. The human liver produces about 600 ml of bile per day.
Portal triad and acinus.
The branches of the portal vein, hepatic artery, and bile duct are located side by side, at the outer border of the lobule, and form the portal triad. There are several such portal triads on the periphery of each lobule.
The functional unit of the liver is the acinus. This is the part of the tissue that surrounds the portal triad and includes the lymphatic vessels, nerve fibers, and adjacent sectors of two or more lobules. One acinus contains about 20 hepatic cells located between the portal triad and the central vein of each lobule. In a two-dimensional image, a simple acinus looks like a group of vessels surrounded by adjacent areas of lobules, and in a three-dimensional image it looks like a berry (acinus - lat. berry) hanging on a stalk of blood and bile vessels. The acinus, whose microvascular frame consists of the above blood and lymphatic vessels, sinusoids and nerves, is the microcirculatory unit of the liver.
liver cells
(hepatocytes) have the shape of polyhedrons, but they have three main functional surfaces: sinusoidal, facing the sinusoidal channel; tubular - participating in the formation of the wall of the bile capillary (it does not have its own wall); and intercellular - directly bordering neighboring liver cells.
Ito cage
Ito cells (synonyms: liver stellate cell, fat-storing cell, lipocyte, English. Hepatic Stellat Cell, HSC, Cell of Ito, Ito cell) - pericytes contained in the perisinusoidal space of the hepatic lobule, capable of functioning in two different states - calm and activated. Activated Ito cells play a major role in fibrogenesis - the formation of scar tissue in liver damage.
In an intact liver, stellate cells are found in calm state. In this state, the cells have several outgrowths covering the sinusoidal capillary. Another distinguishing feature of cells is the presence in their cytoplasm of reserves of vitamin A (retinoid) in the form of fat droplets. Quiet Ito cells make up 5-8% of all liver cells.
Outgrowths of Ito cells are divided into two types: perisinusoidal(subendothelial) and interhepatocellular. The former leave the cell body and extend along the surface of the sinusoidal capillary, covering it with thin finger-like branches. Perisinusoidal outgrowths are covered with short villi and have characteristic long microprotrusions extending even further along the surface of the capillary endothelial tube. Interhepatocellular outgrowths, having overcome the plate of hepatocytes and reaching the neighboring sinusoid, are divided into several perisinusoidal outgrowths. Thus, the Ito cell covers, on average, slightly more than two adjacent sinusoids.
When the liver is damaged, Ito cells become activated state. The activated phenotype is characterized by proliferation, chemotaxis, contractility, loss of retinoid stores, and formation of myofibroblastic-like cells. Activated liver stellate cells also show increased levels of new genes such as α-SMA, ICAM-1, chemokines and cytokines. Activation indicates the beginning of an early stage of fibrogenesis and precedes the increased production of ECM proteins. The final stage of liver healing is characterized by increased apoptosis of activated Ito cells, as a result of which their number is sharply reduced.
To visualize Ito cells under microscopy, staining with gold chloride is used. It was also established that a reliable marker for the differentiation of these cells from other myofibroblasts is their expression of the reelin protein.
Story
In 1876 Karl von Kupfer described the cells he named "Sternzellen" (stellate cells). When stained with gold oxide, inclusions were visible in the cytoplasm of the cells. Mistakenly considering them to be fragments of erythrocytes captured by phagocytosis, Kupfer in 1898 revised his views on the "stellate cell" as a separate type of cell and classified them as phagocytes. However, in subsequent years, descriptions of cells similar to Kupffer's "stellate cells" appeared regularly. They were given various names: interstitial cells, parasinusoid cells, lipocytes, pericytes. The role of these cells remained a mystery for 75 years, until Professor Toshio Ito discovered some cells containing blotches of fat in the perisinusoidal space of the human liver. Ito called them "shibo-sesshu saibo" - fat-absorbing cells. Realizing that the inclusions were fat produced by cells from glycogen, he changed the name to "shibo-chozo saibo" - fat-storing cells. In 1971, Kenjiro Wake proved the identity of Kupffer's "Sternzellen" and Ito's fat-storing cells. Wake also found that these cells play an important role in storing vitamin A (until then it was believed that vitamin A is deposited in Kupffer cells). Shortly thereafter, Kent and Popper demonstrated a close association of Ito cells with liver fibrosis. These discoveries initiated a process of detailed study of Ito cells.
see also
Write a review on the article "Ito's Cage"
Links
- Young-O Queon, Zachary D. Goodman, Jules L. Dienstag, Eugene R. Schiff, Nathaniel A. Brown, Elmar Burckhardt, Robert Skunkhoven, David A. Brenner, Michael W. Fried (2001) . Journal of Haepothology 35; 749-755. - translation of an article in the journal "Infections and Antimicrobial Therapy", Volume 04/N 3/2002, on the Consilium-Medicum website.
- Popper H: Distribution of vitamin A in tissue as revealed by fluorescence microscopy. Physiol Rev 1944, 24:.
Notes
- Geerts A. (2001) History, heterogeneity, developmental biology, and functions of quiescent hepatic stellate cells. Semin Liver Dis. 21(3):311-35. PMID
- Wake, K. (1988) Liver perivascular cells revealed by gold- and silver-impregnation method and electron microscopy. In Biopathology of the Liver. An Ultrastructural Approach” (Motta, P. M., ed) pp. 23-36, Kluwer Academic Publishers, Dordrecht, Netherlands
- Stanciu A, Cotutiu C, Amalinei C. (2002) New data about ITO cells. Rev Med Chir Soc Med Nat Iasi. 107(2):235-9. PMID
- John P. Iredale (2001) Hepatic Stellate Cell Behavior During Resolution of Liver Injury. Seminars in Liver Disease, 21(3):PMID- on Medscape.
- Kobold D, Grundmann A, Piscaglia F, Eisenbach C, Neubauer K, Steffgen J, Ramadori G, Knittel T. (2002) Expression of reelin in hepatic stellate cells and during hepatic tissue repair: a novel marker for the differentiation of HSC from other liver myofibroblasts. J Hepatol. 36(5):607-13. PMID
- Adrian Reuben (2002) hepatology. Volume 35, Issue 2, Pages 503-504
- Suematsu M, Aiso S. (2001) Professor Toshio Ito: a clairvoyant in pericyte biology. Keio J Med. 50(2):66-71. PMID
- Querner F: Der mikroskopische Nachweis von Vitamin A im animalen Gewebe. Zur Kenntnis der paraplasmatischen Leberzellen-einschlüsse. Dritte Mitteilung. Klin Wschr 1935, 14:.
An excerpt characterizing Ito's Cage
Half an hour later, Kutuzov left for Tatarinov, and Bennigsen, with his retinue, including Pierre, rode along the line.
Benigsen descended from Gorki along the high road to the bridge, to which the officer from the mound pointed out to Pierre as the center of the position, and near which rows of mowed grass, smelling of hay, lay on the bank. They drove across the bridge to the village of Borodino, from there they turned left and past a huge number of troops and guns drove to a high mound on which the militias were digging the ground. It was a redoubt, which did not yet have a name, then it was called the Raevsky redoubt, or barrow battery.
Pierre did not pay much attention to this redoubt. He did not know that this place would be more memorable for him than all the places in the Borodino field. Then they drove across the ravine to Semyonovsky, where the soldiers were pulling away the last logs of huts and barns. Then, downhill and uphill, they drove forward through the broken rye, knocked out like hail, along the road to the flushes [a kind of fortification. (Note by L.N. Tolstoy.) ], also then still dug.
Bennigsen stopped at the fleches and began to look ahead at the Shevardinsky redoubt (which had been ours yesterday), on which several horsemen could be seen. The officers said that Napoleon or Murat was there. And everyone looked eagerly at this bunch of riders. Pierre also looked there, trying to guess which of these barely visible people was Napoleon. Finally, the horsemen drove off the mound and disappeared.
Benigsen turned to the general who approached him and began to explain the whole position of our troops. Pierre listened to Benigsen's words, straining all his mental powers to understand the essence of the upcoming battle, but felt with chagrin that his mental abilities were insufficient for this. He didn't understand anything. Bennigsen stopped talking, and noticing the figure of Pierre listening, he suddenly said, turning to him:
- You, I think, are not interested?
“Oh, on the contrary, it’s very interesting,” Pierre repeated, not quite truthfully.
From the flush, they drove even more to the left along the road, winding through a dense, low birch forest. In the middle of it
forest, a brown hare with white legs jumped out in front of them on the road and, frightened by the clatter of a large number of horses, was so confused that he jumped for a long time along the road in front of them, arousing general attention and laughter, and only when several voices shouted at him, rushed to the side and hid in the thicket. Having traveled two versts through the forest, they drove out to a clearing on which stood the troops of Tuchkov's corps, which was supposed to protect the left flank.
Here, on the extreme left flank, Bennigsen spoke a lot and ardently and made, as it seemed to Pierre, an important order from a military point of view. Ahead of the disposition of Tuchkov's troops was an elevation. This elevation was not occupied by troops. Bennigsen loudly criticized this mistake, saying that it was foolish to leave the high ground unoccupied and place troops under it. Some generals expressed the same opinion. One in particular spoke with military vehemence that they were put here to be slaughtered. Bennigsen ordered in his name to move the troops to the heights.
This order on the left flank made Pierre even more doubtful of his ability to understand military affairs. Listening to Bennigsen and the generals who condemned the position of the troops under the mountain, Pierre fully understood them and shared their opinion; but precisely because of this, he could not understand how the one who placed them here under the mountain could make such an obvious and gross mistake.
Pierre did not know that these troops were not sent to defend the position, as Benigsen thought, but were placed in a hidden place for an ambush, that is, in order to be unnoticed and suddenly strike at the advancing enemy. Bennigsen did not know this and moved the troops forward for special reasons, without telling the commander-in-chief about it.
On this clear August evening on the 25th, Prince Andrey was lying, leaning on his arm, in a broken barn in the village of Knyazkov, on the edge of his regiment. Through the hole in the broken wall, he looked at the strip of thirty-year-old birch trees with the lower branches cut off along the fence, at the arable land with smashed heaps of oats on it, and at the bushes, along which smokes of bonfires could be seen - soldiers' kitchens.
No matter how cramped and no one needs and no matter how hard his life now seemed to Prince Andrei, he, just like seven years ago in Austerlitz on the eve of the battle, felt agitated and irritated.
Orders for tomorrow's battle were given and received by him. There was nothing more for him to do. But the simplest, clearest and therefore terrible thoughts did not leave him alone. He knew that tomorrow's battle was to be the most terrible of all those in which he participated, and the possibility of death for the first time in his life, without any relation to worldly things, without considerations of how it would affect others, but only in relation to himself, to his soul, with liveliness, almost with certainty, simply and terribly, she presented herself to him. And from the height of this idea, everything that had previously tormented and occupied him was suddenly illuminated by a cold white light, without shadows, without perspective, without distinction of outlines. All life seemed to him like a magic lantern, into which he looked for a long time through glass and under artificial light. Now he suddenly saw, without glass, in bright daylight, these badly painted pictures. “Yes, yes, here they are, those false images that agitated and delighted and tormented me,” he said to himself, turning over in his imagination the main pictures of his magic lantern of life, now looking at them in this cold white light of day - a clear thought of death. - Here they are, these roughly painted figures, which seemed to be something beautiful and mysterious. Glory, public good, love for a woman, the fatherland itself - how great these pictures seemed to me, what deep meaning they seemed to be filled with! And it's all so simple, pale and crude in the cold white light of that morning that I feel is rising for me." The three main sorrows of his life in particular caught his attention. His love for a woman, the death of his father and the French invasion that captured half of Russia. "Love. This girl, who seemed to me full of mysterious powers. How I loved her! I made poetic plans about love, about happiness with her. O dear boy! he said out loud angrily. - How! I believed in some kind of ideal love, which was supposed to keep her faithful to me during the whole year of my absence! Like the gentle dove of a fable, she must have withered away from me. And all this is much simpler ... All this is terribly simple, disgusting!
Intercellular communication might be realized by paracrine secretion and direct cell-to-cell contacts. It is known that hepatic perisinusoidal cells (HPC) establish regional stem cells niche and determine their differentiation. At the same time HPC remain poorly characterized on the molecular and cellular level.
The aim of the project was to study interactions between rat hepatic perisinusoidal cells and various stem cells such as mononuclear cell fraction of human umbilical cord blood (UCB-MC) and rat bone-marrow derived multipotential mesenchymal stromal cells (BM-MMSC).
materials and methods. Rat BM-MSC and HPC, human UCB-MC cells were derived using standard techniques. To study HPC paracrine regulation we co-cultured UCB-MC or BM-MMSC cells with HPC using Boyden chambers and conditioned HPC cells media. Differentially labeled cells were co-cultured and their interactions were observed by phase-contrast fluorescent microscopy and immunocytochemistry.
results. During the first week of cultivation there was autofluorescence of vitamin A because of fat-storing ability of PHC. BM-MMSC demonstrated high viability in all co-culture models. After 2 day incubation in conditioned media co-culture of BM-MMSC with HPC we observed changes in morphology of MMSC - they decreased in size and their sprouts became shorter. The expression of α-Smooth Muscle Actin and desmin was similar to myofibroblast - an intermediate form of Ito cells culture in vitro. These changes could be due to paracrine stimulation by HPC. The most profound effect of HPC on UCB-MC cells was observed in contact co-culture, thereby it is important for UCB-MC cells to create direct cell-to-cell contacts for maintaining their viability. We did not observe any cell fusion between HPC /UCB and HPC /BM-MMSC cells in co-cultures. In our further experiments we plan to study growth factors produced by HPC for hepatic differentiation of stem cells.
Introduction.
Of particular interest among the variety of liver cells are perisinusoidal liver cells (Ito cells). Due to the secretion of growth factors and extracellular matrix components, they create a microenvironment of hepatocytes, and a number of scientific studies have shown the ability of liver stellate cells to form a microenvironment for progenitor cells (including hematopoietic ones) and influence their differentiation into hepatocytes. Intercellular interactions of these cell populations can be carried out by paracrine secretion of growth factors or direct intercellular contacts, however, the molecular and cellular basis of these processes remain unexplored.
Purpose of the study.
Study of interaction mechanisms Ito cells with hematopoietic (HSC) and mesenchymal (MMSC) stem cells under in vitro conditions.
Materials and methods.
Rat liver Ito cells were isolated by two different enzymatic methods. At the same time, stromal MMSCs were obtained from the bone marrow of rats. Mononuclear fraction of hematopoietic stem cells isolated from human umbilical cord blood. The paracrine effects of Ito cells were studied by culturing MMSCs and HSCs in the medium in which Ito cells grew, and by co-culturing cells separated by a semipermeable membrane. The influence of intercellular contacts has been studied in co-cultivation of cells. For better visualization, each population was labeled with an individual fluorescent label. Cell morphology was assessed by phase-contrast and fluorescence microscopy. Phenotypic features of cultured cells were studied by immunocytochemical analysis.
Results.
Within a week after the isolation of perisinusoidal cells, we noted their ability to autofluorescence due to their fat-accumulating ability. Then the cells passed into an intermediate phase of their growth and acquired a stellate shape. At the initial stages of co-cultivation of Ito cells with rat bone marrow MMSCs, the viability of MMSCs was maintained in all cultivation variants. On the second day, during the cultivation of MMSCs in the culture medium of Ito cells, a change in the morphology of MMSCs occurred - they decreased in size, and the processes shortened. The expression of alpha-smooth muscle actin and desmin in MMSC increased, indicating their phenotypic similarity with myofibroblasts, an intermediate stage of growth of activated Ito cells in vitro. Our data indicate the effect of paracrine factors secreted by Ito cells on the properties of MMSCs in culture.
Based on the co-cultivation of hematopoietic stem cells with Ito cells, it has been shown that hematopoietic stem cells remain viable only when contact co-cultivation with Ito cells. According to the fluorescent analysis of mixed cultures, the phenomenon of fusion of cells from different populations was not revealed.
Conclusions. To maintain the viability of hematopoietic stem cells, the presence of direct intercellular contacts with Ito cells is a decisive factor. Paracrine regulation was noted only when MMSCs were cultivated in a nutrient medium in which Ito cells grew. The study of the influence of specific factors produced by Ito cells on the differentiation of HSCs and MMSCs in cell culture is planned to be carried out in future studies.
Shafigullina A.K., Trondin A.A., Shaikhutdinova A.R., Kaligin M.S., Gazizov I.M., Rizvanov A.A., Gumerova A.A., Kiyasov A.P.
SEI HPE "Kazan State Medical University of the Federal Agency for Health and Social Development"
Genes & Cells: Volume V, No. 1, 2010, pages: 33-40
The authors
Gumerova A.A., Kiyasov A.P.
Regenerative medicine is one of the most rapidly developing and promising areas of medicine, which is based on a fundamentally new approach to the restoration of a damaged organ by stimulating and (or) using stem (progenitor) cells to accelerate the regeneration. In order to put this approach into practice, it is necessary to know what stem cells, and in particular regional stem cells, are, what their phenotype and potency are. For a number of tissues and organs, such as the epidermis and skeletal muscle, stem cells have already been identified and their niches described. However, the liver, an organ whose regenerative abilities have been known since ancient times, has not yet revealed its main secret - the secret of the stem cell. In this review, based on our own and literature data, we discuss the hypothesis put forward that perisinusoidal stellate cells can claim the role of a liver stem cell.
Perisinusoidal liver cells (Ito cells, stellate cells, lipocytes, fat-storing cells, vitamin-A-storing cells) are one of the most mysterious cell types of the liver. The history of the study of these cells dates back more than 130 years, and there are still many more questions regarding their phenotype and functions than answers. The cells were described in 1876 by Kupffer, named by him stellate cells and assigned to macrophages. Later, true sedentary liver macrophages received the name of Kupffer.
It is generally accepted that Ito cells are located in the space of Disse in direct contact with hepatocytes, accumulate vitamin A and are able to produce macromolecules of intercellular substance, and also, having contractile activity, regulate blood flow in sinusoidal capillaries like pericytes. The gold standard for the identification of Ito cells in animals is the identification of the cytoskeletal intermediate filament protein in them, characteristic of muscle tissue - desmin. Other fairly common markers of these cells are markers of neuronal differentiation - acid glial fibrillary protein (Glial fibrillary acid protein, GFAP) and nestin.
For many years, Ito cells were considered only from the standpoint of their participation in the development of fibrosis and cirrhosis of the liver. This is due to the fact that when the liver is damaged, these cells always become activated, which consists in increased expression of desmin, proliferation and transdifferentiation into myofibroblasts-like cell transformation expressing --smooth muscle actin (--GMA) and synthesizing significant amounts of intercellular substance, in particular type I collagen. It is the activity of such activated Ito cells that, according to many researchers, leads to the development of fibrosis and cirrhosis of the liver.
On the other hand, facts are gradually accumulating that allow one to look at Ito cells from completely unexpected positions, namely, as the most important component of the microenvironment for the development of hepatocytes, cholangiocytes and blood cells during the hepatic stage of hematopoiesis, and, moreover, as possible stem ( progenitor) liver cells. The purpose of this review is to analyze current data and views on the nature and functional significance of these cells with an assessment of their possible belonging to the population of liver stem (progenitor) cells.
Ito cells are an important participant in the recovery of the parenchyma during liver regeneration due to the macromolecules of the extracellular matrix produced by them and its remodeling, as well as the production of growth factors. The first doubts about the validity of the established theory, considering Ito cells exclusively as the main culprits of liver fibrosis, appeared when it was found that these cells produce a significant number of morphogenic cytokines. Among them, a significant group is made up of cytokines, which are potential mitogens for hepatocytes.
The most important in this group is hepatocyte growth factor - hepatocyte mitogen, necessary for cell proliferation, survival and motility (it is also known as a scattering factor - scatter factor. A defect in this growth factor and (or) its C-met receptor in mice leads to liver hypoplasia and destruction of its parenchyma as a result of suppression of hepatoblast proliferation, increased apoptosis and insufficient cell adhesion.
In addition to hepatocyte growth factor, Ito cells produce stem cell factor. This has been shown in a model of liver regeneration after partial hepatectomy and exposure to 2-acetoaminofluorene. It has also been found that Ito cells secrete transforming growth factor-- and epidermal growth factor, which play an important role both in the proliferation of hepatocytes during regeneration and stimulate mitosis of the Ito cells themselves. The proliferation of hepatocytes is also triggered by the mesenchymal morphogenic protein epimorphin expressed by Ito cells, which appears in them after partial hepatectomy, and pleiotrophin.
In addition to the paracrine mechanisms of interaction between hepatocytes and Ito cells, direct intercellular contacts of these cells with hepatocytes also play a certain role. The importance of intercellular contacts between Ito cells and epithelial progenitor cells was shown in vitro, when cultivation in mixed culture was more effective for differentiation of the latter into albumin-producing hepatocytes than cultivation of cells separated by a membrane, when they could only exchange soluble factors through cultural environment. Isolated from the fetal liver of a mouse for 13.5 days. mesenchymal cells with the phenotype Thy-1 +/C049!±/vimentin+/desmin+/ --GMA+ after the establishment of direct intercellular contacts stimulated the differentiation of the population of primitive hepatic endodermal cells - into hepatocytes (containing glycogen, expressing mRNA of tyrosine aminotransferase and tryptophanoxyge -names). The population of Thy-1+/desmin+ mesenchymal cells did not express markers of hepatocytes, endothelium, and Kupffer cells, and, most likely, was represented by Ito cells. A high density of desmin-positive Ito cells and their location in close contact with differentiating hepatocytes have been noted in vivo in rat and human prenatal livers. Thus, all these facts allow us to conclude that this cell type is the most important component of the microenvironment, necessary for the normal development of hepatocytes in ontogeny and their recovery in the process of reparative regeneration.
In recent years, data have been obtained indicating a significant effect of Ito cells on the differentiation of hematopoietic stem cells. Thus, Ito cells produce erythropoietin and neurotrophin, which affect the differentiation of not only liver epithelial cells, but also hematopoietic stem cells. The study of fetal hematopoiesis in rats and humans has shown that it is these cells that form the microenvironment of hematopoietic islands in the liver. Ito cells express the vascular cell adhesion molecule-1 (VCAM-1), a key molecule for maintaining adhesion of hematopoietic progenitors to bone marrow stromal cells. In addition, they also express stromal factor-1 - (Stromal derived factor-1 -, SDF-1 -) - a potential chemoattractant for hematopoietic stem cells, stimulating their migration to the site of hematopoiesis due to interaction with the specific receptor Cystein-X- Cystein receptor 4 (CXR4), as well as the homeobox protein Hlx, in the case of a defect in which both the development of the liver itself and hepatic hematopoiesis are disturbed. Most likely, it is the expression of VCAM-1 and SDF-1 a on fetal Ito cells that triggers the recruitment of hematopoietic progenitor cells to the fetal liver for further differentiation. Retinoids accumulated by Ito cells are also an important morphogenesis factor for hematopoietic cells and epithelia. It is impossible not to mention the effect of Ito cells on mesenchymal stem cells. Ito cells isolated from rat liver and fully activated modulate the differentiation of mesenchymal stem cells (multipotent mesenchymal stromal cells) in the bone marrow into hepatocyte-like cells (accumulating glycogen and expressing tetase and phosphoenolpyruvate carboxykinase) after 2 weeks. co-cultivation.
Thus, the accumulated scientific facts allow us to conclude that Ito cells are one of the most important cell types necessary for the development and regeneration of the liver. It is these cells that create the microenvironment both for fetal hepatic hematopoiesis and for the differentiation of hepatocytes during prenatal development, as well as for the differentiation of epithelial and mesenchymal progenitor cells into hepatocytes under in vitro conditions. Currently, these data are not in doubt and are recognized by all researchers of the liver. What, then, served as the starting point for the emergence of the hypothesis put forward in the title of the article?
First of all, its appearance was facilitated by the detection in the liver of cells expressing simultaneously both epithelial markers of hepatocytes and mesenchymal markers of Ito cells. The first works in this area were carried out in the study of prenatal histo- and organogenesis of the liver of mammals. It is the process of development that is the key event, the study of which makes it possible to trace in natural conditions the dynamics of the primary formation of the definitive phenotype of various cell types of an organ using specific markers. Currently, the range of such markers is quite wide. In the works devoted to the study of this issue, various markers of mesenchymal and epithelial cells, individual cell populations of the liver, and stem (including hematopoietic) cells were used.
In the conducted studies, it was found that desmin-positive Ito cells of rat fetuses are transient on 14-15 days. gestations express epithelial markers characteristic of hepatoblasts such as cytokeratins 8 and 18. On the other hand, hepatoblasts at the same time of development express the cell marker Ito desmin. It was this that made it possible to make an assumption about the existence in the liver during intrauterine development of cells with a transitional phenotype expressing both mesenchymal and epithelial markers, and, therefore, to consider the possibility of developing Ito cells and hepatocytes from the same source and (or) consider these cells as one and the same cell type at different stages of development. Further studies on the study of histogenesis, conducted on the material of the human embryonic liver, showed that for 4-8 weeks. In fetal development of the human liver, Ito cells expressed cytokeratins 18 and 19, which was confirmed by double immunohistochemical staining, and weak positive staining for desmin was noted in hepatoblasts.
However, in a work published in 2000, the authors failed to detect the expression of desmin in hepatoblasts in the liver of mouse fetuses, and E-cadherin and cytokeratins in Ito cells. The authors obtained positive staining for cytokeratins in Ito cells in only a small proportion of cases, which they associated with non-specific cross-reactivity of primary antibodies. The choice of these antibodies causes some bewilderment - antibodies to chicken desmin and bovine cytokeratins 8 and 18 were used in the work.
In addition to desmin and cytokeratins, another mesenchymal marker, the vascular cell adhesion molecule VCAM-1, is a common marker for Ito cells and mouse and rat fetal hepatoblasts. VCAM-1 is a unique surface marker that distinguishes Ito cells from myofibroblasts in the adult rat liver and is also present on several other liver cells of mesenchymal origin, such as endotheliocytes or myogenic cells.
Another evidence in favor of the hypothesis under consideration is the possibility of mesenchymal-epithelial transdifferentiation (conversion) of Ito cells isolated from the liver of adult rats. It should be noted that the literature discusses mainly epithelial-mesenchymal rather than mesenchymal-epithelial transdifferentiation, although both directions are recognized as possible, and often the term "epithelial-mesenchymal transdifferentiation" is used to refer to transdifferentiation in any of the directions. After analyzing the expression profile of mRNA and corresponding proteins in Ito cells isolated from the liver of adult rats after exposure to carbon tetrachloride (CTC), the authors found both mesenchymal and epithelial markers in them. Among the mesenchymal markers, nestin, --GMA, matrix metalloproteinase-2 (Matrix Metalloproteinase-2, MMP-2), and among the epithelial markers, muscle pyruvate kinase (Muscle pyruvate kinase, MRK), characteristic of oval cells, cytokeratin 19, a-FP, E-cadherin, as well as the transcription factor Hepatocyte nuclear factor 4- (HNF-4-), specific for cells that are destined to become hepatocytes. It was also found that in the primary culture of human epithelial hepatic progenitor cells, mRNA expression of Itonestin cell markers occurs, GFAP - epithelial progenitors co-express both epithelial and mesenchymal markers. The possibility of mesenchymal-epithelial transdifferentiation is confirmed by the appearance in Ito cells of Integrin-linked kinase (ILK), an enzyme necessary for such transdifferentiation.
Mesenchymal-epithelial transdifferentiation was also revealed in our in vitro experiments, where an original approach was taken to cultivate a pure population of Ito cells isolated from rat liver until a dense cell monolayer was formed. After that, the cells stopped expressing desmin and other mesenchymal markers, acquired the morphology of epithelial cells, and began to express markers characteristic of hepatocytes, in particular, cytokeratins 8 and 18 . Similar results were also obtained during organotypic cultivation of fetal rat liver.
During the last year, two papers have been published in which Ito cells are considered as a subtype of oval cells, or as their derivatives. Oval cells are small oval-shaped cells with a narrow rim of cytoplasm that appear in the liver in some models of toxic liver injury and are currently considered to be bipotent progenitor cells capable of differentiating into both hepatocytes and cholangiocytes. Based on the fact that the genes expressed by isolated Ito cells coincide with the genes expressed by oval cells, and under certain conditions of cultivation of Ito cells, hepatocytes and bile duct cells appear, the authors tested the hypothesis that Ito cells are a type of oval cells capable of generate hepatocytes to regenerate a damaged liver. Transgenic GFAP-Cre/GFP (Green fluorescent protein) mice were fed a methionine-choline-deficient/ethionine-enriched diet to activate Ito cells and oval cells. Resting Ito cells had a GFAP+ phenotype. After Ito cells were activated by injury or culture, their GFAP expression decreased and they began to express markers of oval and mesenchymal cells. The oval cells disappeared when GFP+ hepatocytes appeared, starting to express albumin and eventually replacing large areas of the hepatic parenchyma. Based on their findings, the authors hypothesized that Ito cells are a subtype of oval cells that differentiate into hepatocytes through a "mesenchymal" phase.
In experiments performed on the same model of activation of oval cells, when the latter were isolated from the liver of rats, it was found that in vitro oval cells express not only the traditional markers 0V-6, BD-1/BD-2 and M2RK and markers extracellular matrix, including collagens, matrix metalloproteinases and tissue inhibitors of metalloproteinases - marker features of Ito cells. After exposure to TGF-pl cells, in addition to growth suppression and morphological changes, there was an increase in the expression of these genes, as well as the desmin and GFAP genes, the appearance of the expression of the Snail transcription factor responsible for epithelial-mesenchymal transdifferentiation, and the cessation of E-cadherin expression, which indicates the possibility of "reverse" transdifferentiation of oval cells into Ito cells.
Since oval cells are traditionally considered as bipotent precursors of both hepatocytes and cholangiocytes, attempts were made to establish the possibility of the existence of transitional forms between epithelial cells of the intrahepatic bile ducts and Ito cells. Thus, it was shown that in normal and damaged liver, small structures of the ductal type stained positively for the Ito cell marker - GMA, however, in the photographs presented in the article, which reflect the results of immunofluorescent staining, it is possible to determine what these actually are - GMA+ ductal structures - bile ducts or blood vessels - is not possible. However, other results have been published indicating the expression of Ito cell markers in cholangiocytes. In the already mentioned work of L. Yang, the expression of the Ito cell marker GFAP by bile duct cells was shown. The protein of intermediate filaments of the cytoskeleton sinemine, which is present in normal liver in Ito cells and vascular cells, appeared in the ductal cells involved in the development of the ductular reaction; it was also expressed in cholangi carcinoma cells. Thus, if there is a lot of evidence regarding the possibility of mutual transdifferentiation of Ito cells and hepatocytes, then with cholangiocytes, such observations are still single and not always unambiguous.
Summing up, we can say that the patterns of expression of mesenchymal and epithelial markers both during histo- and organogenesis of the liver, and under various experimental conditions both in vivo and in vitro indicate the possibility of both mesenchymal-epithelial and epithelial-mesenchial small transitions between Ito cells/oval cells/hepatocytes, and therefore, allow us to consider Ito cells as one of the sources of hepatocyte development. These facts undoubtedly point to the inseparable relationship between these cell types, and also indicates a significant phenotypic plasticity of Ito cells. The phenomenal plasticity of these cells is also evidenced by their expression of a number of neural proteins, such as the already mentioned GFAP, nestin, neurotrophins and receptors for them, the neuronal cell adhesion molecule (Neural cell adhesion molecule, N-CAM), synaptophysin, nerve growth factor (Neural growth factor, NGF), brain-derived neurotrophic factor (BDNF), on the basis of which a number of authors discuss the possibility of developing Ito cells from the neural crest. However, over the past decade, researchers have attracted great attention to another version - namely, the possibility of developing hepatocytes and Ito cells from hematopoietic and mesenchymal stem cells.
The first work in which this possibility was proved was published by V.E. Petersen et al., who showed that hepatocytes can develop from a hematopoietic stem cell. Subsequently, this fact was repeatedly confirmed in the works of other scientists, and a little later, the possibility of differentiation into hepatocytes was also shown for mesenchymal stem cells. How this happens - by fusion of donor cells with recipient liver cells, or by their transdifferentiation - is still not clear. However, we also found that human umbilical cord blood hematopoietic stem cells transplanted into the spleen of rats that underwent partial hepatectomy colonize the liver and are able to differentiate into hepatocytes and sinusoidal liver cells, as evidenced by the presence of human cell markers in these cell types. In addition, we have shown for the first time that preliminary genetic modification of umbilical cord blood cells does not significantly affect their distribution and the possibility of differentiation in the recipient's liver after transplantation. As for the likelihood of developing hepatocytes from hematopoietic stem cells during prenatal histogenesis, although this possibility cannot be completely excluded, it nevertheless seems unlikely, since the morphology, localization and phenotype of these cells differ significantly from those for liver cells. Apparently, if such a pathway exists, it does not play a significant role in the formation of epithelial and sinusoidal cells during ontogeny. The results of recent studies, both in vivo and in vitro, cast doubt on the well-established theory of the development of hepatocytes only from the endodermal epithelium of the foregut, and therefore the assumption arose that the regional stem cell of the liver can be located among its mesenchymal cells. Can Ito cells be such cells?
Given the unique properties of these cells, their phenomenal plasticity, and the existence of cells with a transitional phenotype from Ito cells to hepatocytes, we assume that these cells are the main contenders for this role. Additional arguments in favor of this possibility are that these cells, like hepatocytes, can be formed from hematopoietic stem cells, and they are the only sinusoidal liver cells that are able to express markers of stem (progenitor) cells.
In 2004, it was found that Ito cells can also develop from a hematopoietic stem cell. After transplantation of bone marrow cells of GFP mice, GFP+ cells appeared in the liver of recipient mice expressing the Ito cell marker GFAP, and the processes of these cells penetrated between hepatocytes. In case the liver of the recipient was damaged by CTC, the transplanted cells also expressed blast-like Ito cells. When the fraction of non-parenchymal cells was isolated from the liver of recipient mice, GFP+ cells with lipid drops accounted for 33.4+2.3% of the isolated cells; they expressed desmin and GFAP, and after 7 days. cultivation
On the other hand, transplantation of bone marrow cells leads to the formation of not only Ito cells, but the type I collagen gene, on the basis of which it was concluded that such transplantation contributes to the development of fibrosis. However, there are also works where a decrease in liver fibrosis was demonstrated due to the migration of transplanted cells into fibrous septa and the production by these cells of matrix metalloproteinase-9 (Matrix Metalloproteinase-9, MMP-9), which is one of the most important features of Ito cells. Our preliminary data also showed a decrease in the number of myofibroblasts and a decrease in the level of fibrosis after autotransplantation of the peripheral blood mononuclear fraction in patients with chronic hepatitis with severe liver fibrosis. In addition, as a result of hematopoietic stem cell transplantation, other cell types capable of producing extracellular matrix may appear in the recipient's liver. Thus, in case of liver damage induced by bile duct ligation, transplanted cells of differentiated fibrocytes expressing collagen, and only when cultivated in the presence of TGF-pl, are they differentiate-myofibroblasts, potentially contributing to fibrosis. Thus, the authors associated the risk of liver fibrosis after bone marrow cell transplantation not with Ito cells, but with a “unique population of fibrocytes” . Due to the inconsistency of the data obtained, the discussion turned on one more question - whether Ito cells, which appeared as a result of the differentiation of transplanted hematopoietic stem cells, will contribute to the development of fibrosis, or will they provide full-fledged regeneration of liver tissue and reduction of fibrosis. In recent years, it has become obvious (including from the above data) that the origin of myofibroblasts in the liver can be different - from Ito cells, from portal tract fibroblasts, and even from hepatocytes. It has also been established that myofibroblasts of various origins differ in a number of properties. Thus, activated Ito cells differ from portal tract myofibroblasts in terms of vitamin content, contractile activity, response to cytokines, especially TGF-β, and ability to spontaneous apoptosis. In addition, these cell populations are distinct and, where possible, express the vascular cell adhesion molecule VCAM-1, which is present on Ito cells and absent on myofibroblasts. It is impossible not to say that in addition to the production of extracellular matrix proteins, activated Ito cells also produce matrix metalloproteinases that destroy this matrix. Thus, the role of Ito cells, including those formed from hematopoietic stem cells, in the development of fibrosis is far from being as unambiguous as previously thought. Apparently, they do not so much promote fibrosis as remodel the extracellular matrix in the process of liver repair after damage, thus providing a connective tissue scaffold for the regeneration of liver parenchymal cells.
normal and damaged liver of rats. Rat Ito cells also express another marker of stem (progenitor) cells - CD133, and demonstrate the properties of progenitor cells that are capable of differentiating into various types depending on conditions - 2) upon addition of cytokines facilitating differentiation into endothelial cells, form branched tubular structures with induction of expression of markers endothelial cells - endothelial NO-synthase and vascular endothelial cadherin; 3) when using cytokines that promote the differentiation of stem cells into hepatocytes - into rounded cells expressing hepatocyte markers - FP and albumin. Also, rat Ito cells express 0ct4, which is characteristic of pluripotent stem cells. Interestingly, only a part of the Ito cell population can be isolated by a magnetic sorter using anti-CD133 antibodies; however, after standard (pronase/collagenase) isolation, all plastic-attached cells expressed CD133 and 0kt4. Another marker for progenitor cells, Bcl-2 is expressed by desmin+ cells during prenatal development of the human liver.
Thus, various researchers have shown the possibility of expression by Ito cells of certain markers of stem (progenitor) cells. Moreover, an article has recently been published in which for the first time a hypothesis was put forward that the Disse space formed by basement membrane proteins, endothelial cells and hepatocytes, in which Ito cells are located, can constitute a microenvironment for the latter, acting as a “niche” for stem cells. cells. This is evidenced by several features characteristic of the niche of stem cells and identified in the components of the microenvironment of Ito cells. Thus, cells located in close proximity to the stem must produce soluble factors, as well as carry out direct interactions that keep the stem cell in an undifferentiated state and retain it in a niche, often located on the basement membrane. Indeed, the endothelial cells of the sinusoidal capillaries of the liver synthesize soluble SDF-1, which binds specifically to the Ito cell receptor CXR4 and stimulates the migration of these cells in vitro. This interaction plays a key role in the migration of hematopoietic stem cells to their final niche in the bone marrow during ontogenesis and permanent residence in it, as well as in their mobilization into the peripheral blood. It is logical to assume that such an interaction can play a similar role in the liver, keeping Ito cells in the space of Disse. During the early stages of liver regeneration, increased expression of SDF-1 may also help recruit additional body stem cell compartments. The innervation of niche cells should involve the sympathetic nervous system, which is involved in the regulation of recruitment of hematopoietic stem cells. Noradrenergic signals of the sympathetic nervous system play a critical role in GCSF (Granulocyte colony-stimulating factorl-induced mobilization of hematopoietic stem cells from the bone marrow. The location of nerve endings in the immediate vicinity of Ito cells has been confirmed in several works. It has also been found that in response to sympathetic stimulation Ito cells secrete prostaglandins F2a and D, which activate glycogenolysis in nearby parenchymal cells.These facts suggest that the sympathetic nervous system may have an effect on the Ito cell niche.Another function of the stem cell niche is to maintain a "slow" cell cycle and an undifferentiated state of stem cells. cells. The maintenance of the undifferentiated state of Ito cells under in vitro conditions is facilitated by parenchymal liver cells - when these two populations of cells separated by a membrane are cultivated, the expression of stem cell markers CD133 and 0kt4 is preserved in Ito cells, while in the absence of hepatocytes, Ito cells acquire the phenotype of myofibroblasts and lose stem cell markers. Thus, the expression of stem cell markers is undoubtedly a hallmark of resting Ito cells. It has also been established that the influence of parenchymal cells on Ito cells may be based on the interaction of paracrine factors Wnt and Jag1 synthesized by hepatocytes with the corresponding receptors (Myc, Notchl) on the surface of Ito cells. Wnt/b-catenin and Notch signaling pathways support the ability of stem cells to self-renew by slow symmetrical division without subsequent differentiation. Another important component of the niche is the basement membrane proteins, laminin and collagen IV, which maintain the resting state of Ito cells and suppress their differentiation. A similar situation occurs in muscle fibers and convoluted seminiferous tubules, where satellite cells (stem cells of muscle tissue) and undifferentiated spermatogonia are in close contact with the basement membrane, respectively, of the muscle fiber or "spermatogenic epithelium". Obviously, the interaction of stem cells with extracellular matrix proteins inhibits the triggering of their final differentiation. Thus, the data obtained allow us to consider Ito cells as stem cells, a niche for which the space of Disse can serve.
Our data on the stem potency of Ito cells and the possibility of hepatocyte formation from these cells were confirmed in experiments on the study of liver regeneration in vivo in models of partial hepatectomy and toxic damage to the liver with lead nitrate. It is traditionally believed that in these models of liver regeneration there is no activation of the stem compartment and oval cells are absent. We managed to establish, however, that in both cases it is possible to observe not only the activation of Ito cells, but also the expression in them of another stem cell marker, namely, the receptor for the C-kit stem cell factor. Since C-kit expression was also noted in single hepatocytes (in which it was less intense), mainly located in contact with C-kit-positive Ito cells, it can be assumed that these hepatocytes differentiated from C-kit+ Ito cells. It is obvious that this cell type not only creates conditions for the restoration of the hepatocyte population, but also occupies a niche of stem regional liver cells.
Thus, it is now established that Ito cells express at least five stem cell markers under various conditions of development, regeneration and cultivation. All the data accumulated to date suggest that Ito cells can play the role of regional liver stem cells, being one of the sources for the development of hepatocytes (and possibly cholangiocytes), and are also the most important component of the microenvironment for liver morphogenesis and hematopoiesis. Nevertheless, it seems premature to draw unambiguous conclusions about the belonging of these cells to the population of stem (progenitor) cells of the liver. However, there is an obvious need for new research in this direction, which, if successful, will open up prospects for the development of effective methods of treating liver diseases based on stem cell transplantation.