A farkasok izomtevékenységének biokémiája olvasható. A testben zajló biokémiai folyamatok dinamikája izommunka során. összehúzódások, a vázizmok általános áttekintése)
Küldje el a jó munkát a tudásbázis egyszerű. Használja az alábbi űrlapot
Diákok, végzős hallgatók, fiatal tudósok, akik a tudásbázist tanulmányaikban és munkájukban használják, nagyon hálásak lesznek Önnek.
közzétett http://www.allbest.ru/
Bevezetés
1. Vázizmok, izomfehérjék és biokémiai folyamatok az izmokban
2. Biokémiai változások a küzdősportolók szervezetében
4. A gyógyulás problémája a sportban
5. A metabolikus állapotok jellemzői emberben az izomtevékenység során
6. Biokémiai kontroll a harcművészetekben
Következtetés
Bibliográfia
Bevezetés
A biokémia szerepe a modern sportgyakorlatban egyre növekszik. Az izomtevékenység biokémiájának és a testmozgás során az anyagcsere-szabályozás mechanizmusainak ismerete nélkül lehetetlen az edzési folyamat hatékony menedzselése és további racionalizálása. A biokémia ismerete szükséges a sportoló edzettségi szintjének felméréséhez, a túlterhelések és túlerőltetés azonosításához, valamint az étrend helyes megszervezéséhez. A biokémia egyik legfontosabb feladata, hogy a kémiai átalakulások mélyreható ismeretére alapozva hatékony módszereket találjon az anyagcsere szabályozására, hiszen az anyagcsere állapota határozza meg a normalitást és a patológiát. Az élő szervezet növekedése és fejlődése, a külső hatásokkal szembeni ellenálló képessége és az új létfeltételekhez való aktív alkalmazkodás az anyagcsere folyamatok természetétől és sebességétől függ.
Az anyagcsere adaptív változásainak tanulmányozása lehetővé teszi számunkra, hogy jobban megértsük a szervezet fizikai aktivitáshoz való alkalmazkodásának jellemzőit, és hatékony eszközöket és módszereket találjunk a fizikai teljesítmény növelésére.
A küzdősportokban mindig is a fizikai erőnlét problémáját tartották az egyik legfontosabbnak, amely meghatározza a sportteljesítmények szintjét.
Az edzésmódszerek meghatározásának szokásos megközelítése a sportedzés jelenségeit formálisan leíró empirikus törvényszerűségeken alapul.
Maguk a fizikai tulajdonságok azonban nem létezhetnek önmagukban. Úgy jelennek meg, hogy a központi idegrendszer szabályozza az összehúzódó és anyagcsere-energiát pazarló izmokat.
Az elméleti megközelítés megköveteli a sportolói testmodell megalkotását, figyelembe véve a világ sportbiológiájának eredményeit. Az emberi szervezet szerveinek egyes sejtjeiben zajló adaptációs folyamatok irányításához ismerni kell a szerv felépítését, működésének mechanizmusait, valamint azokat a tényezőket, amelyek biztosítják az adaptációs folyamatok célirányát.
1. Vázizmok, izomfehérjék és biokémiai folyamatok az izmokban
A vázizmok nagy mennyiségű nem fehérjeanyagot tartalmaznak, amelyek a fehérje kicsapódása után a zúzott izmokból könnyen vizes oldatba kerülnek. Az ATP közvetlen energiaforrás nemcsak a különféle élettani funkciókhoz (izomösszehúzódások, idegi aktivitás, idegi gerjesztés átvitele, szekréciós folyamatok stb.), hanem a szervezetben lezajló plasztikus folyamatokhoz is (szöveti fehérjék felépítése és megújulása, biológiai szintézisek). Az élet e két aspektusa – az élettani funkciók energiaellátása és a képlékeny folyamatok energiaellátása – között folyamatos a verseny. Rendkívül nehéz bizonyos szabványos normákat megadni azokra a biokémiai változásokra, amelyek egy sportoló szervezetében az egyik vagy másik sport gyakorlása során fellépnek. Az egyes gyakorlatok tiszta formájukban (atlétika futás, korcsolya, síelés) végzett gyakorlatok során is jelentősen eltérhet az anyagcsere folyamatok lefolyása a különböző sportolók között idegi aktivitásuk típusától, környezeti hatásoktól stb. függően. víz és 20-25% szárazanyag. A száraz maradék 85%-a fehérje; a fennmaradó 15%-ot különböző nitrogéntartalmú és nitrogénmentes extraktumok, foszforvegyületek, lipoidok és ásványi sók teszik ki. Izomfehérjék. A szarkoplazmatikus fehérjék az összes izomfehérje 30%-át teszik ki.
Az izomrostok fehérjék az összes izomfehérje körülbelül 40%-át teszik ki. Az izomrostok fehérjéi elsősorban két fő fehérjét foglalnak magukban - a miozint és az aktint. A miozin egy globulin típusú fehérje, amelynek molekulatömege körülbelül 420 000. Sok glutaminsavat, lizint és leucint tartalmaz. Ezenkívül más aminosavakkal együtt ciszteint tartalmaz, ezért szabad csoportokat tartalmaz - SH. A miozin izomrostokban található, az „A korong” vastag filamentumaiban, és nem kaotikusan, hanem szigorúan rendezve. A miozin molekulák fonalas (fibrilláris) szerkezettel rendelkeznek. Huxley szerint hosszuk körülbelül 1500 A, vastagságuk körülbelül 20 A. Az egyik végén vastagság van (40 A). Molekuláinak ezek a végei az „M zónából” mindkét irányban irányulnak, és a vastag filamentumok folyamataiból klub alakú megvastagodásokat képeznek. A miozin a kontraktilis komplex nélkülözhetetlen összetevője, ugyanakkor enzimatikus (adenozin-trifoszfatáz) aktivitással rendelkezik, katalizálja az adenozin-trifoszforsav (ATP) ADP-re és ortofoszfátra történő lebomlását. Az aktin molekulatömege sokkal kisebb, mint a miozin (75 000), és két formában létezhet - globuláris (G-aktin) és fibrilláris (F-aktin) formájában, amelyek képesek egymásba átalakulni. Az első molekulái kerek alakúak; a második molekula, amely a G-aktin polimerje (több molekula kombinációja), fonalas. A G-aktin viszkozitása alacsony, az F-aktin viszkozitása magas. Az aktin egyik formájának a másikba való átmenetét számos ion, különösen K+ és Mg++ segíti elő. Az izomműködés során a G-aktin átalakul F-aktinná. Ez utóbbi könnyen egyesül a miozinnal, és egy aktomiozin nevű komplexet képez, és az izom kontraktilis szubsztrátja, amely képes mechanikai munkát végezni. Az izomfibrillumokban az aktin a „J korong” vékony filamentumaiban helyezkedik el, az „A korong” felső és alsó harmadába nyúlva, ahol az aktin a miozinhoz kapcsolódik a vékony és vastag filamentumok folyamatai közötti érintkezéseken keresztül. A miozin és aktin mellett néhány más fehérje is megtalálható a miofibrillumokban, különösen a vízben oldódó tropomiozin fehérje, amely különösen nagy mennyiségben fordul elő a simaizomzatban és az embriók izomzatában. A fibrillák más, vízben oldódó fehérjéket is tartalmaznak, amelyek enzimatikus aktivitással rendelkeznek” (adenilsav-deamináz stb.). A mitokondriumok és riboszómák fehérjéi főként enzimfehérjék. Különösen a mitokondriumok tartalmaznak aerob oxidációs és légúti foszforilációs enzimeket, a riboszómák pedig fehérjéhez kötött rRNS-t. Az izomrostmagok fehérjéi olyan nukleoproteinek, amelyek molekuláiban dezoxiribonukleinsavakat tartalmaznak.
Az izomrost-sztróma fehérjéi, amelyek az összes izomfehérje körülbelül 20%-át teszik ki. A stromális fehérjékből, az A.Ya. Danilevsky myostromins, felépítette a szarkolemmát és látszólag „Z-korongokat”, amelyek vékony aktinszálakat kapcsoltak össze a szarkolemmával. Lehetséges, hogy a miosztrominokat az aktinnal együtt vékony „J-korongok”-szálai tartalmazzák. Az ATP közvetlen energiaforrás nemcsak a különféle élettani funkciókhoz (izomösszehúzódások, idegi aktivitás, idegi gerjesztés átvitele, szekréciós folyamatok stb.), hanem a szervezetben lezajló plasztikus folyamatokhoz is (szöveti fehérjék felépítése és megújulása, biológiai szintézisek). Az élet e két aspektusa – az élettani funkciók energiaellátása és a képlékeny folyamatok energiaellátása – között folyamatos a verseny. A specifikus funkcionális aktivitás növekedése mindig az ATP-felhasználás növekedésével jár együtt, és ennek következtében csökken a biológiai szintézisekhez való felhasználásának lehetősége. Mint ismeretes, a test szöveteiben, így az izmokban is, fehérjéik folyamatosan megújulnak, de a lebontási és szintézis folyamatai szigorúan kiegyensúlyozottak, a fehérjetartalom szintje állandó marad. Az izomtevékenység során a fehérje megújulás gátolt, és minél többet, annál inkább csökken az ATP-tartalom az izmokban. Következésképpen a maximális és szubmaximális intenzitású edzések során, amikor az ATP újraszintézis túlnyomórészt anaerob módon és legkevésbé teljes mértékben megy végbe, a fehérje megújulás jelentősebben gátolt, mint az átlagos és közepes intenzitású munkavégzés során, amikor az energetikailag nagyon hatékony légúti foszforilációs folyamatok dominálnak. A fehérjemegújulás gátlása az ATP hiányának következménye, amely mind a lebomlási folyamathoz, mind (főleg) a szintézisük folyamatához szükséges. Ezért intenzív izomtevékenység során a fehérjék lebontása és szintézise közötti egyensúly megbomlik, az előbbi dominál az utóbbival szemben. Az izom fehérjetartalma enyhén csökken, a polipeptidek és a nem fehérje jellegű nitrogéntartalmú anyagok tartalma pedig nő. Ezen anyagok egy része, valamint néhány kis molekulatömegű fehérje az izmokat a vérbe hagyja, ahol ennek megfelelően nő a fehérje és a nem fehérje nitrogén tartalma. Ebben az esetben fehérje is megjelenhet a vizeletben. Mindezek a változások különösen jelentősek a nagy intenzitású erőgyakorlatok során. Intenzív izomtevékenység esetén az ammónia képződése is fokozódik az adenozin-monofoszforsav egy részének dezaminációja következtében, amelynek nincs ideje újraszintetizálódni ATP-vé, valamint az ammónia glutaminból történő lehasadása miatt, ami fokozódik az izmok megnövekedett szervetlen foszfáttartalmának hatása, aktiválva a glutamináz enzimet. Növekszik az ammóniatartalom az izmokban és a vérben. A keletkező ammónia eliminációja főként kétféleképpen történhet: az ammónia glutaminsavval történő megkötése glutamin képződéséhez vagy karbamid képződése. Azonban mindkét folyamat megköveteli az ATP részvételét, ezért (tartalmának csökkenése miatt) nehézségekbe ütközik az intenzív izomtevékenység során. Közepes és közepes intenzitású izomtevékenység során, amikor a légúti foszforiláció következtében ATP-reszintézis megy végbe, az ammónia eliminációja jelentősen fokozódik. Csökken a vérben és a szövetekben lévő tartalma, fokozódik a glutamin és a karbamid képződése. A maximális és szubmaximális intenzitású izomtevékenység során az ATP hiánya miatt számos más biológiai szintézis is gátolt. Különösen az acetilkolin szintézise a motoros idegvégződésekben, ami negatívan befolyásolja az idegi gerjesztés átvitelét az izmokhoz.
2. Biokémiai változások a harcművészek testében
A test (dolgozó izmok) energiaszükségletét, mint ismeretes, két fő módon - anaerob és aerob módon - elégítik ki. Ennek a két energiatermelési útnak az aránya a különböző gyakorlatokban változik. Bármely gyakorlat végrehajtása során gyakorlatilag mindhárom energiarendszer működik: az anaerob foszfagén (alaktát) és a tejsav (glikolitikus) és az aerob (oxigén, oxidatív) hatásuk „zónái” részben átfedik egymást. Ezért nehéz elkülöníteni az egyes energiarendszerek „nettó” hozzájárulását, különösen akkor, ha viszonylag rövid ideig működnek. Ebben a tekintetben a „szomszédos” rendszerek energiateljesítményét (működési területét) tekintve gyakran párosítva, foszfagén tejsavval, tejsav oxigénnel. Elsőként az a rendszer jelenik meg, amelynek energiahozzájárulása nagyobb. Az anaerob és aerob energiarendszer relatív terhelése szerint minden gyakorlat anaerob és aerob gyakorlatokra osztható. Az első - túlsúlyban az anaerob, a második - az energiatermelés aerob komponense. Az anaerob gyakorlatok végzésekor vezető minőség az erő (sebesség-erő képességek), az aerob gyakorlatok végzésekor - az állóképesség. A különböző energiatermelő rendszerek aránya nagymértékben meghatározza a különböző fiziológiás rendszerek tevékenységében bekövetkezett változások jellegét és mértékét, amelyek biztosítják a különböző gyakorlatok elvégzését.
Az anaerob gyakorlatoknak három csoportja van: - maximális anaerob teljesítmény (anaerob erő); - közel a maximális anaerob teljesítmény; - szubmaximális anaerob teljesítmény (anaerob-aerob teljesítmény). A maximális anaerob erő (anaerob erő) gyakorlatok olyan gyakorlatok, amelyek szinte kizárólag anaerob módon biztosítják a dolgozó izmok energiaellátását: a teljes energiatermelés anaerob összetevője 90-100%. Főleg a foszfagén energiarendszer (ATP + CP) biztosítja, a tejsav (glikolitikus) rendszer némi részvételével. A kiemelkedő sportolók által a sprint során kifejlesztett rekord maximális anaerob teljesítmény eléri a 120 kcal/perc értéket. Az ilyen gyakorlatok lehetséges maximális időtartama néhány másodperc. A vegetatív rendszerek aktivitásának erősítése a munka során fokozatosan történik. Az anaerob gyakorlatok rövid időtartama miatt végrehajtásuk során a vérkeringés és a légzés funkcióinak nincs ideje elérni a lehetséges maximumukat. A maximális anaerob gyakorlat során a sportoló vagy egyáltalán nem lélegzik, vagy csak néhány légzési ciklust sikerül teljesítenie. Ennek megfelelően az „átlagos” pulmonalis lélegeztetés nem haladja meg a maximum 20-30%-át. A pulzusszám már a rajt előtt is megemelkedik (140-150 ütés/percig), és tovább emelkedik az edzés során, és közvetlenül a cél után éri el legmagasabb értékét - a maximum 80-90%-át (160-180 ütés/perc).
Mivel ezeknek a gyakorlatoknak az energiaalapja az anaerob folyamatok, a szív-légzés (oxigéntranszport) rendszer aktivitásának erősítésének magának a gyakorlatnak az energiaellátása szempontjából gyakorlatilag nincs jelentősége. A laktát koncentrációja a vérben munka közben nagyon keveset változik, bár a dolgozó izmokban elérheti a 10 mmol/kg-ot vagy még többet is a munka végén. A vér laktátkoncentrációja a munka leállítása után néhány percig tovább növekszik, és eléri az 5-8 mmol/l-t. Az anaerob gyakorlatok elvégzése előtt a glükóz koncentrációja a vérben kissé megemelkedik. Megvalósításuk előtt és eredményeként a katekolaminok (adrenalin és noradrenalin) és a növekedési hormon koncentrációja a vérben nagyon jelentősen megnő, de az inzulin koncentrációja kissé csökken; a glukagon és a kortizol koncentrációja nem változik észrevehetően. Ezeknél a gyakorlatoknál a sporteredményeket meghatározó vezető élettani rendszerek és mechanizmusok az izomtevékenység központi idegrendszeri szabályozása (a mozgások összehangolása a nagy izomerő megnyilvánulásával), a neuromuszkuláris rendszer funkcionális tulajdonságai (sebesség-erő), a kapacitás, ill. a dolgozó izmok foszfagén energiarendszerének ereje.
A maximális anaerob erőhöz közeli gyakorlatok (vegyes anaerob erő) olyan gyakorlatok, amelyek túlnyomórészt anaerob energiával látják el a dolgozó izmokat. Az anaerob komponens a teljes energiatermelésben 75-85% - részben a foszfagénnek, nagyobb részben a tejsav (glikolitikus) energiarendszereknek köszönhetően. Az ilyen gyakorlatok lehetséges maximális időtartama kiemelkedő sportolók számára 20-50 másodperc. Ahhoz, hogy ezekhez a gyakorlatokhoz energiát biztosítsunk, az oxigénszállító rendszer aktivitásának jelentős növekedése már bizonyos energetikai szerepet tölt be, és minél nagyobb, minél hosszabb a gyakorlat.
Az edzés során a pulmonalis szellőztetés rohamosan növekszik, így a kb. 1 percig tartó gyakorlat végére elérheti az adott sportoló maximális üzemi szellőztetésének 50-60%-át (60-80 l/perc). A laktát koncentrációja a vérben edzés után nagyon magas - kvalifikált sportolóknál akár 15 mmol/l is lehet. A laktát vérben történő felhalmozódása a dolgozó izmokban nagyon magas képződésével jár (intenzív anaerob glikolízis eredményeként). A glükóz koncentrációja a vérben kissé megemelkedik a nyugalmi állapotokhoz képest (akár 100-120 mg%). A vér hormonális változásai hasonlóak azokhoz, amelyek a maximális anaerob erőterhelés során jelentkeznek.
A maximális anaerob erőhöz közeli gyakorlatok atlétikai teljesítményét meghatározó vezető élettani rendszerek és mechanizmusok megegyeznek az előző csoport gyakorlataival, és ezen felül a dolgozó izmok tejsav (glikolitikus) energiarendszerének ereje. A szubmaximális anaerob erő gyakorlatai (anaerob-aerob erő) olyan gyakorlatok, amelyekben a dolgozó izmok energiaellátásának anaerob összetevője van túlsúlyban. A szervezet teljes energiatermelésében eléri a 60-70%-ot, és főként a tejsav (glikolitikus) energiarendszer biztosítja. Ezen gyakorlatok energiaellátásának jelentős része az oxigén (oxidatív, aerob) energiarendszerhez tartozik. A versenygyakorlatok lehetséges maximális időtartama kiemelkedő sportolók számára 1-2 perc. E gyakorlatok ereje és maximális időtartama olyan, hogy végrehajtásuk során a teljesítménymutatókat. Az oxigénszállító rendszer (pulzusszám, perctérfogat, PV, O2 fogyasztási ráta) megközelítheti vagy akár elérheti is az adott sportolóra vonatkozó maximális értékeket. Minél hosszabb a gyakorlat, annál magasabbak ezek a mutatók a célegyenesben, és annál nagyobb az aerob energiatermelés aránya az edzés során. Ezen gyakorlatok után a dolgozó izmokban és a vérben nagyon magas laktátkoncentrációt rögzítenek - akár 20-25 mmol/l-ig. Így a küzdősportolók edzése és versenytevékenysége körülbelül a sportolók izomzatának maximális terhelésével történik. Ugyanakkor a szervezetben lezajló energiafolyamatokra jellemző, hogy az anaerob gyakorlatok rövid időtartama miatt azok végrehajtása során a vérkeringés és a légzés funkcióinak nincs ideje elérni a lehetséges maximumot. A maximális anaerob gyakorlat során a sportoló vagy egyáltalán nem lélegzik, vagy csak néhány légzési ciklust sikerül teljesítenie. Ennek megfelelően az „átlagos” pulmonalis lélegeztetés nem haladja meg a maximum 20-30%-át.
Egy személy fizikai gyakorlatokat végez, és energiát költ a neuromuszkuláris rendszer segítségével. A neuromuszkuláris rendszer motoros egységek gyűjteménye. Minden motoros egység tartalmaz egy motoros neuront, egy axont és egy sor izomrostot. Az MU mennyisége az emberben változatlan marad. Az MV mennyisége egy izomban lehetséges és az edzés során változtatható, de legfeljebb 5%. Ezért ennek a tényezőnek az izomfunkció növekedésében nincs gyakorlati jelentősége. A CF-en belül számos organellum hiperpláziája (elemszámnövekedése) lép fel: myofibrillumok, mitokondriumok, szarkoplazmatikus retikulum (SRR), glikogéngömbök, mioglobin, riboszómák, DNS stb. A CF-et kiszolgáló kapillárisok száma is változik. A myofibrill az izomrost (sejt) speciális organellumja. Minden állatban megközelítőleg azonos keresztmetszettel rendelkezik. Sorba kapcsolt szarkomerekből áll, amelyek mindegyike aktin- és miozinszálakat tartalmaz. Az aktin és a miozin filamentumok között hidak alakulhatnak ki, és az ATP-ben lévő energiafelhasználással a hidak foroghatnak, pl. myofibrill kontrakció, izomrostok összehúzódása, izomösszehúzódás. A hidak kalciumionok és ATP-molekulák jelenlétében jönnek létre a szarkoplazmában. A myofibrillumok számának növekedése egy izomrostban az erejének, összehúzódási sebességének és méretének növekedéséhez vezet. A myofibrillumok növekedésével párhuzamosan más, a myofibrillumot kiszolgáló organellumok is növekednek, például a szarkoplazmatikus retikulum. A szarkoplazmatikus retikulum belső membránok hálózata, amely hólyagokat, tubulusokat és ciszternákat képez. Az MV-ben az SPR ciszternákat képez, ezekben a ciszternákban halmozódnak fel a kalciumionok (Ca). Feltételezhető, hogy az SPR membránokhoz glikolitikus enzimek kapcsolódnak, ezért az oxigén hozzáférésének leállításakor a csatornák jelentős duzzadását okozzák. Ez a jelenség a hidrogénionok (H) felhalmozódásával függ össze, amelyek a fehérjeszerkezetek részleges tönkremenetelét (denaturációját) és a fehérjemolekulák gyököihez való víz hozzáadását okozzák. Az izomösszehúzódás mechanizmusa szempontjából alapvető fontosságú a Ca szarkoplazmából való kiszivattyúzási sebessége, mivel ez biztosítja az izomrelaxáció folyamatát. Az SPR membránokba nátrium-, kálium- és kalciumpumpák vannak beépítve, így feltételezhető, hogy az SPR membránok felszínének a miofibrillumok tömegéhez viszonyított növekedése az MV relaxáció sebességének növekedéséhez vezet.
Következésképpen az izomrelaxáció maximális sebességének vagy sebességének növekedése (az izom elektromos aktiválásának végétől az izom mechanikai feszültségének nullára csökkenésének időtartama) az SPR membránjainak relatív növekedését jelzi. A maximális tempó fenntartását az MV-ben lévő ATP, KrF tartalékok, a myofibrilláris mitokondriumok tömege, a szarkoplazmatikus mitokondriumok tömege, a glikolitikus enzimek tömege, valamint az izomrost- és vértartalom pufferkapacitása biztosítja.
Mindezek a tényezők befolyásolják az izomösszehúzódás energiaellátásának folyamatát, azonban a maximális tempó fenntartásának képessége elsősorban az SPR mitokondriumától függ. Az oxidatív MV mennyiségének vagy más szóval az izom aerob kapacitásának növelésével a maximális erőnléttel végzett gyakorlatok időtartama megnő. Ez annak köszönhető, hogy a CrF koncentrációjának fenntartása a glikolízis során az MV elsavasodásához, az ATP-fogyasztási folyamatok gátlásához vezet a H-ionok Ca-ionokkal való versengése miatt a miozinfejek aktív központjaiban. Ezért a CrF-koncentráció fenntartásának folyamata az izomban zajló aerob folyamatok túlsúlyával az edzés végrehajtásával egyre hatékonyabbá válik. Fontos az is, hogy a mitokondriumok aktívan szívják fel a hidrogénionokat, ezért rövid távú extrém gyakorlatok (10-30 s) végzése során szerepük inkább a sejtsavasodás pufferelésére korlátozódik. Így az izommunkához való alkalmazkodás a sportoló minden egyes sejtjének munkáján keresztül valósul meg, a sejt élettartama alatti energia-anyagcsere alapján. Ennek a folyamatnak az alapja az ATP fogyasztása a hidrogén- és kalciumionok kölcsönhatása során.
A küzdelmek szórakoztató értékének növelése a verekedés aktivitásának jelentős növekedésével jár együtt a végrehajtott technikai akciók számának egyidejű növekedésével. Ezt figyelembe véve valódi probléma merül fel azzal a ténnyel kapcsolatban, hogy a versenyszerű mérkőzés fokozott intenzitásával a progresszív fizikai fáradtság hátterében a sportoló motoros képességeinek átmeneti automatizálása következik be.
A sportgyakorlatban ez általában egy nagy intenzitással lebonyolított küzdelmes mérkőzés második felében nyilvánul meg. Ebben az esetben (különösen, ha a sportoló nem rendelkezik túl magas szintű speciális állóképességgel) a vér pH-jában jelentős változások figyelhetők meg (7,0 hagyományos egység alatt), ami a sportoló rendkívül kedvezőtlen reakcióját jelzi az ilyen intenzitású munkára. Ismeretes, hogy például a birkózó motoros képességeinek ritmikus szerkezetének stabil megzavarása hátrahajlás dobása során a fizikai fáradtság szintjével kezdődik 7,2 arb alatti vér pH-értékeknél. egységek
Ebben a tekintetben kétféle módon lehet növelni a harcművészek motorikus képességeinek stabilitását: a) a speciális állóképesség szintjének olyan szintre emelése, hogy bármilyen intenzitású küzdelmet le tudjanak vívni kifejezett fizikai fáradtság nélkül (a reakció a terhelés nem vezethet acidotikus eltolódáshoz a 7,2 hagyományos egységgel megegyező pH-érték alá); b) biztosítsa a motoros készségek stabil megnyilvánulását extrém fizikai aktivitás bármilyen extrém helyzetében a 6,9-es hagyományos értéket elérő vér pH-értékeknél. egységek Az első irány keretében meglehetősen nagy számú speciális tanulmányt végeztek, amelyek meghatározták a harcművészeti sportolók speciális állóképességének felgyorsított edzésének problémájának megoldásának valódi módjait és kilátásait. A második probléma kapcsán a mai napig nem történt valódi, gyakorlatilag jelentős fejlemény.
4. A gyógyulás problémája a sportban
Az edzési folyamat intenzívebbé tételének és a sportteljesítmény további növelésének egyik legfontosabb feltétele a helyreállító eszközök széleskörű és szisztematikus alkalmazása. A racionális felépülés különösen fontos az extrém és a maximumhoz közeli fizikai és mentális stressz során – ez az edzés és a versenyek kötelező szatellitei a modern sportokban. Nyilvánvaló, hogy a helyreállító eszközök rendszerének alkalmazása szükségessé teszi a helyreállítási folyamatok egyértelmű osztályozását a sporttevékenység körülményei között.
A felépülési változások sajátossága, amelyet a sporttevékenység jellege, az edzés és a versenyterhelések mennyisége és intenzitása, valamint az általános rezsim határoz meg, meghatározza a teljesítmény helyreállítását célzó konkrét intézkedéseket. N. I. Volkov a következő típusú gyógyulást azonosítja a sportolóknál: aktuális (munka közbeni megfigyelés), sürgős (a terhelés végét követő) és késleltetett (a munka befejezése után több órával), valamint krónikus túlerőltetés után (ún. stressz helyreállítása). Meg kell jegyezni, hogy a felsorolt reakciók a normál életkörülmények közötti energiafogyasztás miatti időszakos helyreállítás hátterében zajlanak.
Jellegét nagymértékben meghatározza a szervezet funkcionális állapota. A felépülési folyamatok dinamikájának világos megértése a sporttevékenység körülményei között szükséges a helyreállítási eszközök ésszerű használatának megszervezéséhez. Így a folyamatos felépülés során kialakuló funkcionális változások a szervezet fokozott energiaszükségletének biztosítását, az izomtevékenység folyamatában fellépő megnövekedett biológiai energiafogyasztás kompenzálását célozzák. Az anyagcsere-átalakítások központi helyet foglalnak el az energiaköltségek helyreállításában.
A szervezet energiafelhasználásának és munka közbeni helyreállításának aránya lehetővé teszi a fizikai aktivitás 3 tartományba történő felosztását: 1) terhelések, amelyeknél elegendő a munka aerob támogatása; 2) olyan terhelések, amelyekben a munka aerob támogatásával együtt anaerob energiaforrásokat használnak, de a dolgozó izmok oxigénellátásának növelésének határát még nem lépték túl; 3) olyan terhelések, amelyeknél az energiaszükséglet meghaladja az áram-visszanyerési képességeket, amihez gyorsan fejlődő fáradtság társul. Egyes sportágakban a rehabilitációs intézkedések hatékonyságának felméréséhez tanácsos a neuromuszkuláris rendszer különféle mutatóinak elemzése és pszichológiai tesztek alkalmazása. A magas színvonalú sportolókkal végzett munka gyakorlati alkalmazása széles körű eszközök és módszerek segítségével lehetővé teszi számunkra, hogy értékeljük a korábbi rehabilitációs intézkedések hatékonyságát, és meghatározzuk a későbbiek taktikáját. A felépülési tesztek szakaszos vizsgálatokat igényelnek heti vagy havi edzési ciklusokban. E vizsgálatok és kutatási módszerek gyakoriságát az orvos és az edző határozza meg a sportág típusától, az adott edzési időszak terheléseinek jellegétől, az alkalmazott helyreállító eszközöktől és a sportoló egyéni jellemzőitől függően.
5 . A metabolikus állapotok jellemzői emberben az izomtevékenység során
Az emberi szervezet anyagcseréjének állapotát nagyszámú változó jellemzi. Intenzív izomtevékenység körülményei között a legfontosabb tényező, amelytől a szervezet anyagcsere-állapota függ, az energia-anyagcsere területén való alkalmazás. Az emberi anyagcsere-állapotok számszerűsítésére az izommunka során háromféle kritérium alkalmazását javasoljuk: a) teljesítménykritériumok, amelyek az aerob és anaerob folyamatok energiakonverziójának sebességét tükrözik; b) a szervezet energiatartalékait vagy a munka során bekövetkezett anyagcsere-változások teljes volumenét jellemző kapacitáskritériumokat; c) hatékonysági kritériumok, amelyek meghatározzák, hogy az aerob és anaerob folyamatok energiája milyen mértékben kerül felhasználásra izommunka végzése során. Az edzés teljesítményének és időtartamának változása eltérő hatással van az aerob és az anaerob anyagcserére. Az aerob folyamat teljesítményének és kapacitásának olyan mutatói, mint a pulmonalis lélegeztetés mérete, az oxigénfogyasztás szintje és a munka közbeni oxigénbevitel, szisztematikusan nőnek az edzés időtartamával minden kiválasztott teljesítményérték mellett. Ezek a mutatók észrevehetően nőnek a munka intenzitásának növekedésével a gyakorlat minden időintervallumában. Az anaerob energiaforrások kapacitását jellemző tejsav maximális felhalmozódása a vérben és a teljes oxigéntartozás mutatói mérsékelt erejű gyakorlatok végzése során keveset változnak, intenzívebb gyakorlatok esetén viszont érezhetően nőnek a munkaidő növekedésével.
Érdekes megjegyezni, hogy a legalacsonyabb terhelés mellett, ahol a vér tejsavtartalma körülbelül 50-60 mg állandó szinten marad, gyakorlatilag lehetetlen kimutatni az oxigéntartozás laktátfrakcióját; A tejsav felhalmozódása során a vér bikarbonátjainak megsemmisüléséhez kapcsolódó szén-dioxid túlzott felszabadulása nem következik be. Feltételezhető, hogy a tejsav felhalmozódásának jelzett szintje a vérben még nem haladja meg azokat a küszöbértékeket, amelyek felett a laktát-oxigén-tartozás megszüntetésével járó oxidatív folyamatok stimulálása figyelhető meg. Az aerob anyagcsere indikátorai az edzéssel kapcsolatos rövid (kb. 1 perces) késleltetési időszak után szisztémás növekedést mutatnak az edzésidő növekedésével.
A bejáratási időszakban az anaerob reakciók jelentős növekedése következik be, ami tejsav képződéséhez vezet. Az edzésteljesítmény növekedése az aerob folyamatok arányos növekedésével jár együtt. Az aerob folyamatok intenzitásának növekedését növekvő teljesítménnyel csak azokban a gyakorlatokban állapították meg, amelyek időtartama meghaladja a 0,5 percet. Intenzív, rövid távú gyakorlatok végzésekor az aerob anyagcsere csökkenése figyelhető meg. A teljes oxigéntartozás növekedése a laktátfrakció képződése és a többlet szén-dioxid felszabadulás megjelenése miatt csak azoknál a gyakorlatoknál észlelhető, amelyek ereje és időtartama elegendő ahhoz, hogy 50-60 mg tejsav halmozódjon fel. %. Kis teljesítményű gyakorlatok végzésekor az aerob és anaerob folyamatok mutatóinak változása ellentétes irányt mutat, az erőnövekedéssel ezek a folyamatok egyirányúvá változnak.
Az oxigénfogyasztás sebességének és a „túlzott” szén-dioxid-felszabadulásnak az edzés közbeni mutatóinak dinamikájában fáziseltolódás észlelhető; a munka befejezését követő helyreállítási időszakban ezeknek a mutatóknak az eltolódásai szinkronizálódnak. Az intenzív edzés utáni felépülési idő növekedésével az oxigénfogyasztás és a vér tejsavszintjének változása egyértelműen fáziskülönbségeket mutat. A fáradtság problémája a sport biokémiájában az egyik legnehezebb, és még mindig messze van a megoldástól. A fáradtság legáltalánosabb formájában a szervezetnek a hosszan tartó vagy megerőltető tevékenység következtében fellépő állapotaként definiálható, és teljesítménycsökkenéssel jellemezhető. Szubjektív módon a személy helyi fáradtság vagy általános fáradtság érzéseként érzékeli. A hosszú távú vizsgálatok lehetővé teszik, hogy a teljesítményt korlátozó biokémiai tényezőket három, egymással összefüggő csoportba soroljuk.
Ezek először is a központi idegrendszer biokémiai változásai, amelyeket mind maga a motoros gerjesztési folyamat, mind a perifériáról érkező proprioceptív impulzusok okoznak. Másodszor, ezek a vázizmok és a szívizom biokémiai változásai, amelyeket munkájuk és az idegrendszer trofikus változásai okoznak. Harmadszor, ezek biokémiai változások a test belső környezetében, mind az izmokban végbemenő folyamatoktól, mind az idegrendszer hatásától függően. A fáradtság közös jellemzője a foszfát makroergek egyensúlyának felborulása az izmokban és az agyban, valamint az ATP-áz aktivitás és a foszforilációs együttható csökkenése az izmokban. A nagy intenzitású és hosszú ideig tartó munkával járó fáradtságnak azonban vannak sajátos jellemzői is. Emellett a rövid távú izomtevékenység okozta fáradtság során fellépő biokémiai változásokat lényegesen nagyobb gradiens jellemzi, mint a közepes intenzitású izomtevékenység során, de időtartama közel van a határhoz. Hangsúlyozni kell, hogy a szervezet szénhidráttartalékainak éles csökkenése, bár nagy jelentőséggel bír, nem játszik döntő szerepet a teljesítmény korlátozásában. A teljesítményt korlátozó legfontosabb tényező az ATP szintje mind magukban az izmokban, mind a központi idegrendszerben.
Ugyanakkor nem lehet figyelmen kívül hagyni a biokémiai változásokat más szervekben, különösen a szívizomban. Intenzív, rövid távú munkával a glikogén és a kreatin-foszfát szintje nem változik, de az oxidatív enzimek aktivitása nő. Hosszan tartó munkavégzés esetén a glikogén és a kreatin-foszfát szintje, valamint az enzimaktivitás is csökkenhet. Ehhez dystrophiás folyamatokat jelző EKG-elváltozások társulnak, leggyakrabban a bal kamrában, ritkábban a pitvarban. Így a fáradtságra mind a központi idegrendszerben, mind a periférián, elsősorban az izmokban mélyreható biokémiai változások jellemzőek. Sőt, az utóbbi biokémiai változásainak mértéke a központi idegrendszerre gyakorolt hatás által okozott teljesítménynövekedéssel változtatható. I.M. 1903-ban írt a fáradtság központi idegrendszeri természetéről. Sechenov. Azóta egyre több adat áll rendelkezésre a központi gátlásnak a fáradtság mechanizmusában betöltött szerepéről. Az elhúzódó izomtevékenység okozta fáradtság során kialakuló diffúz gátlás megléte nem kétséges. A központi idegrendszerben fejlődik ki és fejlődik benne a centrum és a periféria kölcsönhatása révén az előbbiek vezető szerepével. A fáradtság az intenzív vagy hosszan tartó tevékenység által a szervezetben bekövetkezett változások következménye, és egy olyan védőreakció, amely megakadályozza az átmenetet a szervezetre veszélyes és létét veszélyeztető funkcionális és biokémiai rendellenességek vonalán.
A fáradtság mechanizmusában bizonyos szerepet játszanak az idegrendszer fehérje- és nukleinsav-anyagcsere zavarai is. A hosszan tartó, jelentős fáradtságot okozó terheléses futás vagy úszás során a motoros neuronokban RNS-szint csökkenés figyelhető meg, míg hosszan tartó, de nem fárasztó munkavégzés során nem változik, vagy emelkedik. Mivel a kémiát és különösen az izomenzimek aktivitását az idegrendszer trofikus hatásai szabályozzák, feltételezhető, hogy az idegsejtek kémiai állapotának változásai a fáradtság okozta védőgátlás kialakulása során a trofikus centrifugális változásokhoz vezetnek. impulzusok, ami zavarokhoz vezet az izomkémia szabályozásában.
Ezek a trofikus hatások nyilvánvalóan a biológiailag aktív anyagok mozgásán keresztül valósulnak meg az efferens rostok axoplazmájában, ahogy azt P. Weiss leírta. A perifériás idegekből egy fehérjeanyagot izoláltak, amely a hexokináz specifikus inhibitora, hasonlóan ennek az enzimnek az agyalapi mirigy elülső része által kiválasztott inhibitorához. Így a fáradtság a központi és a perifériás mechanizmusok kölcsönhatása révén alakul ki, az előbbiek vezető és integráló fontosságával. Mind az idegsejtek változásaihoz, mind a perifériáról érkező reflex- és humorális hatásokhoz kapcsolódik. A fáradtság során fellépő biokémiai változások általánosíthatók, a szervezet belső környezetének általános változásaival, valamint a különböző élettani funkciók szabályozásának és koordinációjának zavaraival (hosszú, jelentős izomtömeggel járó fizikai aktivitás során). Ezek a változások lehetnek lokálisabb természetűek is, nem kísérik jelentős általános változások, hanem csak a dolgozó izmokra és a megfelelő idegsejtek és központok csoportjaira korlátozódnak (rövid idejű maximális intenzitású vagy hosszú távú korlátozott munkavégzés során). izmok száma).
A fáradtság (és különösen a fáradtság érzése) védőreakció, amely megvédi a szervezetet a túlzott mértékű funkcionális kimerültségtől, ami életveszélyes. Ugyanakkor edzi a fiziológiai és biokémiai kompenzációs mechanizmusokat, megteremtve a felépülési folyamatok előfeltételeit és tovább növelve a szervezet funkcionalitását és teljesítőképességét. Az izommunka utáni pihenés során a biológiai vegyületek normális aránya helyreáll mind az izmokban, mind a test egészében. Ha izommunka során az energiaellátáshoz szükséges katabolikus folyamatok dominálnak, akkor pihenéskor az anabolikus folyamatok dominálnak. Az anabolikus folyamatok energiafelhasználást igényelnek ATP formájában, ezért a legkifejezettebb változások az energia-anyagcsere területén tapasztalhatók, mivel a pihenőidő alatt az ATP folyamatosan elhasználódik, ezért az ATP-tartalékokat helyre kell állítani. A pihenőidő alatti anabolikus folyamatok a munka során fellépő katabolikus folyamatoknak köszönhetőek. A pihenés során az ATP, a kreatin-foszfát, a glikogén, a foszfolipidek és az izomfehérjék újraszintetizálódnak, a szervezet víz-elektrolit egyensúlya visszaáll a normális szintre, és helyreállnak a sérült sejtszerkezetek. A szervezetben végbemenő biokémiai változások általános irányától és az elválasztási folyamatokhoz szükséges időtől függően kétféle helyreállítási folyamatot különböztetünk meg - sürgős és felhagyott gyógyulást. A sürgős gyógyulás a munka után 30-90 percig tart. A sürgős gyógyulás időszakában a munkavégzés során felhalmozódott anaerob bomlástermékek, elsősorban a tejsav- és oxigéntartozás megszűnnek. A munka befejezése után az oxigénfogyasztás továbbra is magasabb a nyugalmi állapothoz képest. Ezt a túlzott oxigénfogyasztást oxigéntartozásnak nevezzük. Az oxigéntartozás mindig nagyobb, mint az oxigénhiány, és minél nagyobb a munka intenzitása és időtartama, annál jelentősebb ez a különbség.
Pihenés közben az izomösszehúzódásokhoz szükséges ATP fogyasztás leáll, és az első másodpercekben megnő a mitokondriumok ATP-tartalma, ami a mitokondriumok aktív állapotba való átmenetét jelzi. Az ATP-koncentráció növekszik, növelve a munka előtti szintet. Az oxidatív enzimek aktivitása is megnő. De a glikogén-foszforiláz aktivitása élesen csökken. A tejsav, mint már tudjuk, a glükóz anaerob körülmények közötti lebomlásának végterméke. A pihenés kezdeti pillanatában, amikor a fokozott oxigénfogyasztás megmarad, az izmok oxidatív rendszereinek oxigénellátása fokozódik. A tejsav mellett a munka során felhalmozódott egyéb metabolitok is oxidációnak vannak kitéve: borostyánkősav, glükóz; a gyógyulás későbbi szakaszaiban pedig a zsírsavak. A késés helyreállítása a munka befejezése után sokáig tart. Elsősorban az izommunka során elhasznált struktúrák szintézisének folyamatait, valamint a szervezet ion- és hormonális egyensúlyának helyreállítását érinti. A gyógyulási időszak alatt a glikogén tartalékok felhalmozódnak az izmokban és a májban; ezek a helyreállítási folyamatok 12-48 órán belül lezajlanak. A vérbe jutó tejsav a májsejtekbe jut, ahol először a glükóz szintézis megy végbe, a glükóz pedig a glikogénszintézist katalizáló glikogén-szintetáz közvetlen építőanyaga. A glikogén újraszintézis folyamata fázisos jellegű, amely a szuperkompenzáció jelenségén alapul. A szuperkompenzáció (túlvisszanyerés) az energiaanyag-tartalékoknak a pihenőidő alatti munkaszintre való túllépése. A szuperkompenzáció elfogadható jelenség. A munka után lecsökkent glikogéntartalom a pihenés során nemcsak a kezdeti szintre, hanem magasabb szintre is emelkedik. Ezután következik a csökkenés a kezdeti (dolgozó) szintre és még egy kicsit lejjebb, majd hullámszerűen visszatér az eredeti szintre.
A szuperkompenzációs fázis időtartama a munka időtartamától és az általa a szervezetben kiváltott biokémiai változások mélységétől függ. Az erőteljes, rövid távú munka a szuperkompenzációs fázis gyors beindulását és gyors befejezését idézi elő: amikor az intramuszkuláris glikogéntartalékok helyreállnak, a szuperkompenzációs fázis 3-4 óra elteltével észlelhető és 12 óra múlva véget ér. Hosszan tartó, közepes teljesítményű munka után a glikogén szuperkompenzációja 12 óra elteltével következik be, és a munka befejezése után 48 és 72 óra között ér véget. A szuperkompenzáció törvénye minden olyan biológiai vegyületre és szerkezetre érvényes, amelyek valamilyen mértékben elfogynak vagy megszakadnak az izomtevékenység során, és pihenés közben újraszintetizálódnak. Ide tartoznak: kreatin-foszfát, szerkezeti és enzimatikus fehérjék, foszfolipidek, sejtorgonellák (mitokondriumok, lizoszómák). A szervezet energiatartalékainak újraszintézise után a foszfolipidek és fehérjék újraszintézisének folyamatai jelentősen felgyorsulnak, különösen nehéz erőmunka után, amely jelentős lebontással jár. A strukturális és enzimatikus fehérjék szintjének helyreállítása 12-72 órán belül megtörténik. Vízvesztéssel járó munkavégzés során a gyógyulási időszak alatt pótolni kell a víz- és ásványi sók tartalékait. Az ásványi sók fő forrása az élelmiszer.
6 . Biokémiai kontroll a harcművészetekben
Az intenzív izomtevékenység során az izmokban nagy mennyiségű tej- és piroszőlősav képződik, amelyek a vérbe diffundálva a szervezet metabolikus acidózisát okozhatják, ami izomfáradtsághoz vezet, és izomfájdalommal, szédüléssel, hányingerrel jár. Az ilyen anyagcsere-változások a szervezet puffertartalékainak kimerülésével járnak. Mivel a szervezet pufferrendszereinek állapota fontos a magas fizikai teljesítmény megnyilvánulásában, a CBS indikátorokat a sportdiagnosztikában használják. A CBS-indikátorok, amelyek általában viszonylag állandóak, a következők: - a vér pH-ja (7,35-7,45); - pCO2 - a szén-dioxid (H2CO3 + CO2) parciális nyomása a vérben (35-45 Hgmm); - 5B - standard vérplazma-hidrogén-karbonát HSOd, amely ha a vér teljesen oxigénnel telített, 22-26 meq/l; - BB - teljes vér vagy plazma pufferbázisai (43 - 53 meq/l) - a vér vagy plazma teljes pufferrendszerének kapacitásának mutatója; - L/86 - a teljes vér normál pufferbázisai az alveoláris levegő pH és CO2 fiziológiás értékein; - BE - bázisfelesleg vagy lúgos tartalék (-2,4 és +2,3 meq/l között) - a puffer feleslegének vagy hiányának mutatója. A CBS indikátorok nemcsak a vérpufferrendszerek változásait tükrözik, hanem a szervezet légző- és kiválasztórendszerének állapotát is. A szervezet sav-bázis egyensúlyának (ABC) állapotát a vér állandó pH-értéke (7,34-7,36) jellemzi.
Fordított korrelációt állapítottak meg a vér laktáttartalmának dinamikája és a vér pH-jának változása között. Az ABS indikátorok izomtevékenység közbeni megváltoztatásával nyomon követhető a szervezet fizikai aktivitásra adott reakciója és a sportoló edzettségének növekedése, mivel az ABS biokémiai szabályozásával ezek közül az indikátorok egyike meghatározható. A vizelet aktív reakciója (pH) közvetlenül függ a szervezet sav-bázis állapotától. Metabolikus acidózis esetén a vizelet savassága pH 5-re emelkedik, metabolikus alkalózis esetén pH 7-re csökken. Táblázat. A 3. ábra a vizelet pH-értékeinek változásának irányát mutatja a plazma sav-bázis állapotának mutatóihoz viszonyítva. Így a birkózást mint sportágat az izomtevékenység nagy intenzitása jellemzi. Ebben a tekintetben fontos ellenőrizni a savak cseréjét a sportoló testében. Az ACS leginformatívabb mutatója a BE - lúgos tartalék értéke, amely a sportolók, különösen a gyorsasági-erős sportokra szakosodott sportolók képzettségének növekedésével növekszik.
Következtetés
Összegzésképpen elmondható, hogy a harcművészek edzése és versenytevékenysége körülbelül a sportolók izomzatának maximális terhelése mellett zajlik. Ugyanakkor a szervezetben lezajló energiafolyamatokra jellemző, hogy az anaerob gyakorlatok rövid időtartama miatt azok végrehajtása során a vérkeringés és a légzés funkcióinak nincs ideje elérni a lehetséges maximumot. A maximális anaerob gyakorlat során a sportoló vagy egyáltalán nem lélegzik, vagy csak néhány légzési ciklust sikerül teljesítenie. Ennek megfelelően az „átlagos” pulmonalis lélegeztetés nem haladja meg a maximum 20-30%-át. A küzdősportolók verseny- és edzéstevékenységében a fáradtság az izmok közel maximális terhelése miatt jelentkezik a küzdelem teljes időtartama alatt.
Ennek eredményeként a vér pH-ja megemelkedik, a sportoló reakciója és az ellenség támadásaival szembeni ellenállása romlik. A fáradtság csökkentése érdekében javasolt glikolitikus anaerob terhelés alkalmazása az edzési folyamatban. A domináns fókusz által létrehozott nyomfolyamat meglehetősen tartós és inert lehet, ami lehetővé teszi a gerjesztés fenntartását még az irritáció forrásának eltávolításakor is.
Az izommunka befejezése után kezdődik a felépülési, vagy utómunkálati időszak. Jellemzője a testfunkciók változásának mértéke és az eredeti szintre való visszaállításukhoz szükséges idő. A helyreállítási időszak tanulmányozása szükséges egy adott munka súlyosságának felméréséhez, annak meghatározásához, hogy megfelel-e a szervezet képességeinek, és meghatározza a szükséges pihenés időtartamát. A harcművészek motorikus készségeinek biokémiai alapja közvetlenül összefügg az erőképességek megnyilvánulásával, amelyek közé tartozik a dinamikus, robbanékony és izometrikus erő. Az izommunkához való alkalmazkodás a sportoló minden egyes sejtjének munkáján keresztül történik, a sejt élettartama alatti energia-anyagcsere alapján. Ennek a folyamatnak az alapja az ATP fogyasztása a hidrogén- és kalciumionok kölcsönhatása során. A harcművészetet, mint sportágat nagy intenzitású izomtevékenység jellemzi. Ebben a tekintetben fontos ellenőrizni a savak cseréjét a sportoló testében. Az ACS leginformatívabb mutatója a BE - lúgos tartalék értéke, amely a sportolók, különösen a gyorsasági-erős sportokra szakosodott sportolók képzettségének növekedésével növekszik.
Bibliográfia
1. Volkov N.I. Az izomtevékenység biokémiája. - M.: Olimpiai sport, 2001.
2. Volkov N.I., Oleynyikov V.I. A sport bioenergiája. - M: Szovjet sport, 2011.
3. Maksimov D.V., Seluyanov V.N., Tabakov S.E. Harcművészek fizikai felkészítése. - M: TVT osztály, 2011.
Közzétéve az Allbest.ru oldalon
Hasonló dokumentumok
A citoplazma mozgásszervi rendszere. Az izomszövet szerkezete és kémiai összetétele. Az izmok funkcionális biokémiája. Bioenergetikai folyamatok az izomtevékenység során. A fizikai gyakorlatok biokémiája. Biokémiai változások az izmokban a patológia során.
képzési kézikönyv, hozzáadva 2009.07.19
A fogalom lényege és az izomtevékenység főbb funkciói. Az emberi test helyreállítási szakasza. A helyreállítás mutatói és a folyamatot felgyorsító eszközök. A gyorskorcsolya főbb élettani jellemzői.
teszt, hozzáadva: 2008.11.30
Az edzési folyamat biokémiai monitorozása. A laboratóriumi ellenőrzés típusai. A szervezet energiaellátó rendszere. A sportolók táplálkozásának jellemzői. Az energiaátalakítás módjai. A képzettség foka, az alkalmazkodás főbb típusai, jellemzőik.
szakdolgozat, hozzáadva: 2018.01.22
Az izmok, mint az emberi test szervei, amelyek idegimpulzusok hatására összehúzódó izomszövetből állnak, osztályozásuk, típusuk, funkcionális szerepük. Az emberi test izommunkájának jellemzői, dinamikus és statikus.
bemutató, hozzáadva 2013.04.23
A vázizom tömege felnőtteknél. A mozgásszervi rendszer aktív része. Harántcsíkolt izomrostok. A vázizmok felépítése, főcsoportjai és simaizomzata és munkájuk. Az izomrendszer életkorral összefüggő jellemzői.
teszt, hozzáadva: 2009.02.19
Biokémiai elemzések a klinikai gyógyászatban. Vérplazma fehérjék. Májbetegségek, gyomor-bél traktus, vérzéscsillapító zavarok, vérszegénység és vérátömlesztés, diabetes mellitus, endokrin betegségek klinikai biokémiája.
képzési kézikönyv, hozzáadva 2009.07.19
A szívizomszövet fejlődési forrásainak jellemzői, amelyek a precordialis mezodermában találhatók. A kardiomiociták differenciálódásának elemzése. A szívizomszövet szerkezetének jellemzői. A szívizomszövet regenerációs folyamatának lényege.
bemutató, hozzáadva 2012.11.07
Biokémiai elemzések a klinikai gyógyászatban. Az univerzális kóros jelenségek patokémiai mechanizmusai. Klinikai biokémia reumás betegségekre, légzőrendszeri, vese- és gyomor-bélrendszeri betegségekre. A hemosztázis rendszer zavarai.
képzési kézikönyv, hozzáadva 2009.07.19
A gyermek testi-lelki fejlődése újszülött- és csecsemőkorban. Az óvodáskor előtti életszakasz anatómiai és élettani sajátosságai. Az izomrendszer és a csontváz fejlesztése általános iskolás korú gyermekeknél. A pubertás időszaka gyermekeknél.
bemutató, hozzáadva 2015.10.03
A jól kialakított és működő mozgásszervi rendszer a gyermek megfelelő fejlődésének egyik fő feltétele. Ismerkedés a gyermekek csont- és izomrendszerének főbb jellemzőivel. Az újszülött mellkasának általános jellemzői.
A sportélettanban szokás megkülönböztetni és felosztani az izomtevékenységet erőzónákra: maximális, szubmaximális, magas és közepes. Az izommunka egy másik felosztása is az energiaellátás fő mechanizmusaitól függ: az anaerob, kevert és aerob energiaellátási zónákban.
Minden izmos munka során mindenekelőtt különbséget kell tenni a kezdeti (kezdő) fázis és a folytatás között. A kezdő fázis ideje a munka intenzitásától függ: minél hosszabb a munka, annál intenzívebb a kezdő szakasz, és annál markánsabbak az izomzatban az alatta végbemenő biokémiai változások.
A munka első másodperceiben az izmok kevesebb oxigént kapnak, mint amennyire szükségük van. Minél nagyobb a munka intenzitása, és ennek megfelelően minél nagyobb az oxigénigény, annál nagyobb az oxigénhiány. Ezért a kiindulási fázisban az ATP újraszintézise kizárólag anaerob módon megy végbe, a kreatin-kináz reakció és a glikolízis következtében.
Ha az izommunka intenzitása maximális és ennek megfelelően rövid az időtartam, akkor az ebben a kezdő fázisban ér véget. Ebben az esetben az oxigénigény nem lesz kielégítve.
Ha szubmaximális intenzitással, de hosszabb ideig dolgozunk, a biokémiai változások a kezdő fázisban kevésbé lesznek drámaiak, és maga a kezdő fázis lerövidül. Ebben az esetben az oxigénfogyasztás eléri az MPC-t (maximális lehetséges értékeket), de az oxigénigény továbbra sem lesz kielégítve. Ilyen körülmények között a szervezet oxigénhiányt tapasztal. Csökken a kreatin-kináz útvonal jelentősége, a glikolízis elég intenzíven fog lezajlani, de az aerob ATP újraszintézis mechanizmusai már aktiválódnak. A glikolízis folyamata főként a vér által a májból származó glükózt foglalja magában, nem pedig az izomglikogénből képződő glükózt.
Még kisebb intenzitású és hosszabb ideig tartó izommunka során egy rövid idejű kezdési fázist követően az aerob mechanizmus révén az ATP újraszintézis dominál, ami az oxigénigény és az oxigénellátás közötti valódi egyensúly megteremtésének a következménye. Az izomrostokban az ATP szintje emelkedik és stabilizálódik, de ez a szint alacsonyabb, mint nyugalmi állapotban. Ezenkívül a kreatin-foszfát szintje enyhén megemelkedik.
Ha hosszan tartó izommunka során ereje élesen megnő, akkor ugyanazok a jelenségek figyelhetők meg, mint a kezdő fázisban. Az üzemi teljesítmény növekedése természetesen az oxigénigény növekedésével jár, amelyet nem lehet azonnal kielégíteni. Ennek eredményeként az ATP újraszintézis anaerob mechanizmusai aktiválódnak.
Tekintsük a különböző ATP újraszintézis utak aktiválásának időbeli sorrendjét. Az izommunka első 2-3 másodpercében energiaellátása az izom ATP lebomlásának köszönhető. 3-20 másodpercig az ATP újraszintézise a kreatin-foszfát lebomlása miatt következik be. Ezután az izommunka kezdete után 30-40 másodperccel a glikolízis eléri a legnagyobb intenzitást. Továbbá az oxidatív foszforiláció folyamatai kezdenek egyre fontosabb szerepet játszani az energiaellátásban (10. ábra).
10. ábra. Különféle energiaforrások részvétele az izomtevékenység energiaellátásában, annak időtartamától függően: 1 - ATP lebontása, 2 - kreatin-foszfát lebontása,
3 – glikolízis, 4 – aerob oxidáció
Az aerob energiatermelés teljesítményét a MIC értékkel becsüljük meg. A statisztikák azt mutatják, hogy a férfiak átlagosan magasabb BMD-vel rendelkeznek, mint a nők. A sportolóknál ez az érték lényegesen magasabb, mint az edzetlen embereknél. A különböző szakterületek sportolói közül a legmagasabb VO2 max értékek a síelők és a hosszútávfutók körében figyelhetők meg.
A szisztematikus fizikai aktivitás az izomsejtekben a mitokondriumok számának növekedéséhez, valamint a légzőlánc enzimek számának és aktivitásának növekedéséhez vezet. Ez megteremti a feltételeket a beérkező oxigén teljesebb felhasználásához és a sikeresebb energiaellátáshoz egy edzett szervezetben.
Rendszeres edzéssel megnő az izmokat vérrel ellátó erek száma. Ez hatékonyabb rendszert hoz létre az izmok oxigénnel és glükózzal való ellátására, valamint a salakanyagok eltávolítására. A hosszú távú edzés során a keringési és légzőrendszer úgy alkalmazkodik, hogy az első gyakorlatok után fellépő oxigéntartozás utólag teljesen kompenzálható. Az izmok hosszú távú munkavégzési képessége általában függ az oxigénfelvétel és -felhasználás sebességétől és hatékonyságától.
Az izomrendszer és funkciói
összehúzódások, a vázizmok általános áttekintése)
Kétféle izom létezik: sima(akaratlan) és barázdált(tetszőleges). A simaizmok az erek falában és egyes belső szervekben találhatók. Összeszűkítik vagy kitágítják az ereket, a táplálékot a gyomor-bél traktuson mozgatják, és összehúzzák a hólyag falát. A harántcsíkolt izmok mind olyan vázizmok, amelyek különféle testmozgásokat biztosítanak. A harántcsíkolt izmok közé tartozik a szívizom is, amely automatikusan biztosítja a szív ritmikus működését egész életen át. Az izmok alapja a fehérjék, amelyek az izomszövet 80-85%-át alkotják (a víz kivételével). Az izomszövet fő tulajdonsága az kontraktilitás, kontraktilis izomfehérjék – aktin és miozin – biztosítják.
Az izomszövet nagyon összetett. Az izomnak rostos szerkezete van, minden rost egy miniatűr izom, ezeknek a rostoknak a kombinációja alkotja az izom egészét. izom rost, viszont abból áll myofibrillumok Mindegyik myofibrill váltakozó világos és sötét területekre oszlik. Sötét területek - a protofibrillumok hosszú molekulaláncokból állnak miozin, a világosakat vékonyabb fehérjeszálak alkotják aktina. Amikor az izom összehúzódás nélküli (lazított) állapotban van, az aktin és a miozin filamentumok csak részben vannak előrehaladva egymáshoz képest, minden egyes miozin filamentum szemben áll, és több aktin filamentum veszi körül. Az egymáshoz viszonyított mélyebb előrehaladás az egyes izomrostok és az egész izom myofibrillumainak rövidülését (összehúzódását) okozza (2.3. ábra).
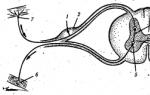
Számos idegrost közeledik és távozik az izomból (reflexív elv) (2.4. ábra). A motoros (efferens) idegrostok impulzusokat továbbítanak az agyból és a gerincvelőből, ezáltal az izmokat működőképes állapotba hozzák; szenzoros rostok az ellenkező irányú impulzusokat továbbítják, tájékoztatva a központi idegrendszert az izomtevékenységről. A szimpatikus idegrostokon keresztül az izomzat anyagcsere-folyamatait szabályozzák, ezáltal tevékenységük alkalmazkodik a megváltozott munkakörülményekhez és a különféle izomterhelésekhez. Minden izmot kiterjedt kapillárishálózat hatol át, amelyen keresztül az izmok működéséhez szükséges anyagok bejutnak és az anyagcseretermékek kiürülnek.
Vázizmok. A vázizmok a váz- és izomrendszer szerkezetének részét képezik, a csontváz csontjaihoz kapcsolódnak, és összehúzódásukkor a váz egyes részeit és a karokat mozgatják. Részt vesznek a test és részei térbeli helyzetének megőrzésében, mozgást biztosítanak járás, futás, rágás, nyelés, légzés stb. során, miközben hőt termelnek. A vázizmok képesek izgatottak lenni az idegimpulzusok hatására. A gerjesztést kontraktilis struktúrákra (miofibrillákra) hajtják végre, amelyek összehúzódva bizonyos motoros aktust - mozgást vagy feszültséget - hajtanak végre.
Rizs. 2.3. Az izom sematikus ábrázolása.
Az izom (L) izomrostokból áll (B), mindegyik myofibrillákból áll (BAN BEN). Myofibrill (G) vastag és vékony myofilamentumokból áll (D). Az ábrán egy szarkomer látható, amelyet mindkét oldalról vonalak határolnak: 1 - izotróp korong, 2 - anizotróp korong, 3 - kisebb anizotrópiával rendelkező terület. Multifibrillák keresztirányú közege (4), képet ad a vastag és vékony multifilamentumok hatszögletű eloszlásáról

Rizs. 2.4. A legegyszerűbb reflexív diagramja:
1 - afferens (érzékeny) neuron, 2 - gerinccsomó, 3 - interneuron, 4 .- a gerincvelő szürkeállománya, 5 - efferens (motoros) neuron, 6 - motoros idegvégződés az izmokban; 7 - szenzoros idegvégződés a bőrben
Emlékezzünk vissza, hogy minden vázizom harántcsíkolt izmokból áll. Az emberekben körülbelül 600 van belőlük, és legtöbbjük páros. Súlyuk a felnőttek teljes testtömegének 35-40%-át teszi ki. A vázizmokat kívülről sűrű kötőszöveti membrán borítja. Minden izomnak van egy aktív része (izomtest) és egy passzív része (ín). Az izmok fel vannak osztva hosszú rövidÉs széles.
Azokat az izmokat, amelyek működése ellentétes irányú, nevezzük antagonisták egyirányú - szinergisták. Ugyanazok az izmok különböző helyzetekben egy és másik minőségben működhetnek. Az embereknél az orsó alakú és a szalag alakúak gyakoribbak. Fusiform izmok A végtagok hosszú csontképződményeinek területén helyezkednek el és működnek, két hasuk (digasztrikus izomzat) és több fejük (bicepsz, tricepsz, négyfejű izomzat) lehet. Szalag izmok különböző szélességűek, és általában részt vesznek a test falainak fűzőképzésében. A tollas szerkezetű izmok, amelyek nagy fiziológiai átmérőjűek a sok rövid izomszerkezet miatt, sokkal erősebbek, mint azok az izmok, amelyekben a rostok lineáris (hosszirányú) elrendezésűek. Az előbbieket erős izmoknak, amelyek kis amplitúdójú mozdulatokat hajtanak végre, az utóbbiakat ügyes izmoknak, amelyek nagy amplitúdójú mozgásokban vesznek részt. Az ízületek funkcionális célja és mozgásiránya szerint az izmokat megkülönböztetik hajlítókÉs extensorok, adduktorokÉs abducens, záróizmok(kompresszív) és bővítők.
Izomerő a teher súlya határozza meg, amelyet egy bizonyos magasságig képes felemelni (vagy maximális gerjesztéssel képes megtartani) anélkül, hogy a hossza megváltozna. Az izom ereje az izomrostok erőinek összegétől és kontraktilitásától függ; az izomrostok számáról és a funkcionális egységek számáról, egyidejűleg izgatott, amikor feszültség alakul ki; tól től kezdeti izomhossz(az előfeszített izomzat nagyobb erőt fejleszt); tól től a vázcsontokkal való interakció feltételei.
Összehúzódás izomra jellemző annak abszolút erő, azok. erő az izomrostok 1 cm 2 keresztmetszetére. Ennek a mutatónak a kiszámításához az izomerőt területtel osztják fiziológiai átmérője(azaz az izmot alkotó összes izomrost területének összege). Például: az átlagember rendelkezik a gastrocnemius izom erejével (1 cm 2 izomkeresztmetszetre vetítve). - 6,24; nyakfeszítők - 9,0; tricepsz brachii izom - 16,8 kg.
A központi idegrendszer szabályozza az izomösszehúzódás erejét azáltal, hogy megváltoztatja az összehúzódásban egyidejűleg részt vevő funkcionális egységek számát, valamint a hozzájuk küldött impulzusok gyakoriságát. Az impulzusfrekvencia növekedése a feszültség növekedéséhez vezet.
Izommunka. Az izomösszehúzódás folyamata során a potenciális kémiai energia potenciális mechanikai feszültségi energiává és mozgási energiává alakul. Különbséget kell tenni a belső és a külső munka között. A belső munkához kapcsolódik az izomrost súrlódása annak összehúzódása során. A külső munka a saját test, a terhelés vagy az egyes testrészek térbeli mozgatásakor nyilvánul meg (dinamikus munka). Jellemzője az izomrendszer hatékonysági tényezője (hatékonysága), i.e. az elvégzett munka aránya a teljes energiafelhasználáshoz (az emberi izomzat hatékonysága 15-20%, a fizikailag fejlett, edzett embereknél valamivel magasabb).
Statikus erőfeszítésekkel (mozgás nélkül) a fizika szempontjából nem a munkáról, mint olyanról beszélhetünk, hanem munkáról, amelyet a szervezet élettani energiaköltségei alapján kell értékelni.
Az izom mint szerv.Általában az izom mint szerv egy összetett szerkezeti képződmény, amely bizonyos funkciókat lát el, és 72-80%-ban vízből és 16-20%-ban sűrű anyagból áll. Az izomrostok sejtmagokkal, riboszómákkal, mitokondriumokkal, szarkoplazmatikus retikulummal, érzékeny idegképződményekkel - proprioceptorokkal és egyéb funkcionális elemekből állnak, amelyek biztosítják a fehérjeszintézist, az adenozin-trifoszforsav oxidatív foszforilációját és reszintézisét, anyagok szállítását az izomsejten belül stb. izomrostok működése során. Az izom fontos szerkezeti és funkcionális képződménye a motoros, vagy neuromotoros egység, amely egy motoros neuronból és az általa beidegzett izomrostokból áll. Az összehúzódásban részt vevő izomrostok számától függően vannak kis, közepes és nagy motoros egységek.
A kötőszöveti rétegekből és membránokból álló rendszer köti össze az izomrostokat egyetlen működő rendszerré, amely az inak segítségével az izomösszehúzódás során fellépő vontatást továbbítja a váz csontjainak.
Az egész izomzatot elágazó erek és nyirokágak hálózata hatol át. balekok. Vörös izomrostok sűrűbb érhálózattal rendelkeznek, mint fehér. Nagy mennyiségű glikogén- és lipidkészlettel rendelkeznek, jelentős tónusos aktivitás jellemzi őket, képesek elviselni a hosszan tartó stresszt és hosszan tartó dinamikus munkát végeznek. Minden vörös rostban több mitokondrium található, mint a fehér rostokban - energiatermelők és -szolgáltatók, 3-5 kapilláris veszi körül, és ez megteremti a feltételeket a vörös rostok intenzívebb vérellátásához és az anyagcsere folyamatok magas szintjéhez.
Fehér izomrostok miofibrillumaik vastagabbak és erősebbek, mint a vörös rostok myofibrillumai, gyorsan összehúzódnak, de nem képesek hosszan tartó feszültségre. A fehérállomány mitokondriumának csak egy kapillárisa van. A legtöbb izom különböző arányban tartalmaz vörös és fehér rostokat. Vannak izomrostok is tonik(terjedése nélkül képes helyi gerjesztésre); fázis,.képes a terjedő gerjesztési hullámra összehúzódással és relaxációval egyaránt reagálni; átmeneti, kombinálva mindkét tulajdonságot.
Izompumpa- az izomműködéssel és annak saját vérellátására gyakorolt hatásával összefüggő élettani fogalom. Fő hatása a következőképpen nyilvánul meg: a vázizmok összehúzódása során az artériás vér beáramlása lelassul, és felgyorsul a vénákon keresztüli kiáramlása; a relaxáció időszakában a vénás kiáramlás csökken, az artériás beáramlás eléri a maximumot. A vér és a szövetfolyadék közötti anyagcsere a kapilláris falán keresztül történik.

Rizs. 2.5. ben lezajló folyamatok sematikus ábrázolása
szinapszis gerjesztéskor:
1 - szinaptikus vezikulák, 2 - preszinaptikus membrán, 3 - közvetítő, 4 - posztszinaptikus membrán, 5 - szinaptikus hasadék
Az izomzat mechanizmusai Az izomműködést különféle szabályozások szabályozzák csökkentések a központi idegrendszer (CNS) osztályai, amelyek nagymértékben meghatározzák sokoldalú tevékenységük jellegét
(mozgás fázisai, tónusos feszültség stb.). Receptorok A motoros apparátusból a motoros analizátor afferens rostjai keletkeznek, amelyek a gerincvelő felé tartó vegyes (afferens-efferens) idegrostok 30-50%-át teszik ki. Izomösszehúzódás Impulzusokat okoz, amelyek az izomérzet forrásai - kinesztézia.
A gerjesztés átvitele az idegrostokról az izomrostokra keresztül történik neuromuszkuláris csomópont(2.5. ábra), amely két réssel elválasztott membránból áll - preszinaptikus (ideg eredetű) és posztszinaptikus (izom eredetű). Idegimpulzus hatására acetilkolin mennyisége szabadul fel, ami egy elektromos potenciál megjelenéséhez vezet, amely gerjesztheti az izomrostokat. Az idegimpulzusok szinapszison keresztüli átvitelének sebessége ezerszer kisebb, mint egy idegrostban. Gerjesztést csak az izom irányába vezet. Normális esetben akár 150 impulzus is áthaladhat az emlős neuromuszkuláris csomópontján egy másodperc alatt. Fáradtsággal (vagy patológiával) a neuromuszkuláris végződések mobilitása csökken, és az impulzusok jellege megváltozhat.
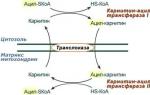
Az izomösszehúzódás kémiája és energiája. Az izom összehúzódása és feszülése a kémiai átalakulások során felszabaduló energia miatt történik, amely az izomba való belépéskor történik.
idegimpulzussal vagy közvetlen irritációval. A kémiai átalakulások az izmokban úgy történnek, mint oxigén jelenlétében(aerob körülmények között) és távollétében(anaerob körülmények között).
Az adenozin-trifoszforsav (ATP) hasítása és újraszintézise. Az izomösszehúzódás elsődleges energiaforrása az ATP (a sejtmembránban, a retikulumban és a miozinszálakban található) lebontása adenozin-difoszforsavra (ADP) és foszforsavakra. Ebben az esetben 10 000 cal szabadul fel minden gramm ATP-molekulából:
ATP = ADP + H3PO4 + 10 000 cal.
A további átalakulások során az ADP adenilsavvá defoszforilálódik. Az ATP lebomlását az aktomiozin fehérje enzim (adenozin-trifoszfatáz) serkenti. Nyugalomban nem aktív; akkor aktiválódik, amikor az izomrost izgatott. Az ATP viszont a miozin filamentumokra hat, növelve azok nyújthatóságát. Az aktomiozin aktivitása megnő a Ca-ionok hatására, amelyek nyugalmi állapotban a szarkoplazmatikus retikulumban helyezkednek el.
Az izomban lévő ATP-tartalékok jelentéktelenek, aktivitásuk fenntartásához folyamatos ATP-reszintézis szükséges. Ez a kreatin-foszfát (CrP) kreatinná (Cr) és foszforsavvá (anaerob fázis) történő lebontásából nyert energia miatt következik be. Az enzimek segítségével a KrP-ből származó foszfátcsoport gyorsan (ezredmásodperceken belül) átkerül az ADP-be. Ebben az esetben minden mól CrP 46 kJ szabadul fel:
És így, az izom teljes energiafelhasználását biztosító végső folyamat az oxidációs folyamat. Eközben a hosszú távú izomműködés csak akkor lehetséges, ha elegendő oxigénellátás van hozzá, hiszen Az energia leadására képes anyagok tartalma anaerob körülmények között fokozatosan csökken. Ezenkívül a tejsav felhalmozódik, a reakció savas oldalra való eltolódása megzavarja az enzimreakciókat, és az anyagcsere gátlásához és dezorganizációjához, valamint az izomteljesítmény csökkenéséhez vezethet. Hasonló állapotok lépnek fel az emberi szervezetben a maximális, szubmaximális és nagy intenzitású (teljesítményű) munka során, például rövid és közepes távok futása során. A kialakult hipoxia (oxigénhiány) miatt az ATP nem áll helyre teljesen, úgynevezett oxigéntartozás keletkezik, tejsav halmozódik fel.
Az ATP aerob újraszintézise(szinonimák: oxidatív foszforiláció, szöveti légzés) - 20-szor hatékonyabb, mint az anaerob energiatermelés. A tejsavnak az anaerob tevékenység során és a hosszan tartó munkavégzés során felhalmozódott része szén-dioxiddá és vízzé oxidálódik (1/4-1/6), a keletkező energiát a maradék tejsavrészek helyreállítására fordítják. glükózba és glikogénbe, miközben biztosítja az ATP és a KrF újraszintézisét. Az oxidatív folyamatok energiáját az izom azonnali működéséhez szükséges szénhidrátok újraszintézisére is felhasználják.
Általában a szénhidrátok adják a legtöbb energiát az izommunkához. Például a glükóz aerob oxidációja során 38 ATP molekula képződik (összehasonlításképpen: a szénhidrát anaerob lebontása során mindössze 2 ATP molekula képződik).
Aerob útvonal kiépítési ideje Az ATP képződése 3-4 perc (edzetteknél - legfeljebb 1 perc), a maximális teljesítmény 350-450 cal/min/kg, a maximális teljesítmény fenntartásának ideje több tíz perc. Ha nyugalomban az ATP aerob újraszintézisének sebessége alacsony, akkor a fizikai aktivitás során az ereje maximális lesz, és ugyanakkor az aerob út órákon át működhet. Rendkívül gazdaságos is: a folyamat során a kiindulási anyagok mélyen lebomlanak a végtermékekké CO2 és NaO. Ezenkívül az ATP-reszintézis aerob útvonalát a szubsztrátok felhasználásának sokoldalúsága jellemzi: a szervezet összes szerves anyaga oxidálódik (aminosavak, fehérjék, szénhidrátok, zsírsavak, ketontestek stb.).
Az ATP-reszintézis aerob módszerének azonban vannak hátrányai is: 1) oxigénfogyasztást igényel, amelynek az izomszövetekbe juttatását a légzőrendszer és a szív- és érrendszer biztosítja, ami természetesen ezek feszültségével jár együtt; 2) a mitokondriális membránok állapotát és tulajdonságait befolyásoló bármely tényező megzavarja az ATP képződését; 3) az aerob ATP képződés kialakulása hosszú ideig tart és alacsony teljesítményű.
A legtöbb sportban végzett izomtevékenységet az ATP újraszintézisének aerob folyamata nem tudja teljes mértékben biztosítani, és a szervezet kénytelen anaerob ATP-képzési módszereket is beiktatni, amelyek rövidebb kifejtési idejűek és nagyobb a folyamat maximális teljesítménye ( azaz az egységnyi idő alatt képződő legnagyobb mennyiségű ATP - 1 mol ATP 7,3 cal-nak vagy 40 J-nak felel meg (1 cal == 4,19 J).
Visszatérve az energiaképzés anaerob folyamataira, tisztázni kell, hogy ezek legalább kétféle reakcióban fordulnak elő: 1. Kreatin-foszfokináz - amikor a CrP lehasad, a foszforcsoportok, amelyekből az ADP-be kerülnek, újraszintetizálva az ATP-t. De az izmokban kicsi a kreatin-foszfát tartalék, és ez az ilyen típusú reakciók gyors (2-4 másodpercen belüli) kihalását okozza. 2. Glikolitikus(glikolízis) - lassabban, intenzív munkavégzés után 2-3 percen belül alakul ki. A glikolízis az izomglikogén tartalékok és a vércukorszint foszforilálásával kezdődik. Ennek a folyamatnak az energiája több percnyi kemény munkához elegendő. Ebben a szakaszban a glikogén-foszforiláció első szakasza befejeződik, és megtörténik az oxidatív folyamatra való felkészülés. Ezután következik a glikolitikus reakció második szakasza - a dehidrogénezés és a harmadik - az ADP redukálása ATP-vé. A glikolitikus reakció két tejsavmolekula képződésével végződik, amely után a légzési folyamatok kibontakoznak (3-5 perces munkavégzéskor), amikor az anaerob reakciók során keletkező tejsav (laktát) oxidálódni kezd.
Az ATP-reszintézis kreatin-foszfát anaerob útvonalának felmérésére szolgáló biokémiai indikátorok a kreatinin együttható és az tejsav (tejsav nélküli) oxigéntartozás. Kreatinin arány- a kreatinin vizelettel történő kiválasztódása naponta 1 testtömegkilogrammonként. Férfiaknál a kreatinin kiválasztódása 18-32 mg/nap x kg, nőknél 10-25 mg/nap x kg. Lineáris kapcsolat van a kreatin-foszfát tartalma és a kreatinin képződése között. Ezért a kreatinin-koefficiens segítségével felmérhető ennek az ATP-reszintézis-útvonalnak a potenciális képességei.
A tejsav felhalmozódása által okozott biokémiai változások a szervezetben glikolízis eredményeként. Ha nyugalomban van a nyaki tevékenység megkezdése előtt laktát koncentráció a vérben 1-2 mmol/l, majd intenzív, rövid ideig tartó, 2-3 perces terhelés után ez az érték elérheti a 18-20 mmol/l-t. Egy másik mutató, amely tükrözi a tejsav felhalmozódását a vérben vérszámlálás(pH): nyugalomban 7,36, edzés után 7,0-ra vagy többre csökken. A laktát vérben való felhalmozódása határozza meg annak lúgos tartalék - az összes vérpufferrendszer lúgos komponensei.
Az intenzív izomtevékenység végét az oxigénfogyasztás csökkenése kíséri – kezdetben élesen, majd fokozatosan. Ezzel kapcsolatban kiemelik az oxigénadósság két összetevője: gyors (alaktát) és lassú (laktát). laktát - ennyi oxigént használnak fel a munka befejezése után a tejsav eltávolítására: kisebb része J-bO-vá és COa-vá oxidálódik, nagyobb része glikogénné alakul. Ez az átalakulás jelentős mennyiségű ATP-t igényel, amely aerob módon képződik az oxigén hatására, amely képződik laktát adósság. A laktát metabolizmusa a májban és a szívizom sejtjeiben megy végbe.
Az elvégzett munka teljes biztosításához szükséges oxigénmennyiséget ún oxigénigény. Például egy 400 m-es versenyen az oxigénigény körülbelül 27 liter. A táv világrekord szinten történő lefutásának ideje körülbelül 40 másodperc. Tanulmányok kimutatták, hogy ezalatt a sportoló 3-4 liter 02-t szív fel. Ezért 24 liter teljes oxigéntartozás(az oxigénigény kb. 90%-a), ami a verseny után megszűnik.
A 100 m-es futamban az oxigéntartozás elérheti a kereslet 96%-át is. A 800 méteres távon az anaerob reakciók aránya kismértékben - 77%-ra, a 10 000 méteres távon - 10%-ra csökken, i.e. az energia túlnyomó részét légúti (aerob) reakciók szolgáltatják.
Az izomlazítás mechanizmusa. Amint az idegimpulzusok abbahagyják az izomrostba jutást, a Ca2-ionok az úgynevezett kalciumpumpa hatására, az ATP energiája miatt, a szarkoplazmatikus retikulum ciszternáiba kerülnek, és koncentrációjuk a szarkoplazmában a kezdeti értékre csökken. szint. Ez változásokat okoz a troponin konformációjában, ami a tropomiozint az aktinszálak egy bizonyos területén rögzítve lehetetlenné teszi a vastag és vékony filamentumok közötti kereszthidak kialakulását. Az izomrostot körülvevő kollagénszálakban az izomösszehúzódás során fellépő rugalmas erők hatására az ellazuláskor visszatér eredeti állapotába. Így az izomlazítás vagy relaxáció folyamata, valamint az izomösszehúzódás folyamata az ATP hidrolízis energiájával történik.
Az izomtevékenység során az összehúzódási és ellazulási folyamatok felváltva mennek végbe az izmokban, ezért az izmok gyorsasági-erőssége egyformán függ az izomösszehúzódás sebességétől és az izmok ellazulási képességétől.
A simaizomrostok rövid jellemzői. A simaizomrostokból hiányoznak a myofibrillumok. Vékony filamentumok (aktin) kapcsolódnak a szarkolemmához, vastag filamentumok (miozin) az izomsejteken belül helyezkednek el. A simaizomrostokból szintén hiányoznak a Ca-ionokat tartalmazó ciszternák. Idegimpulzus hatására a Ca-ionok lassan belépnek a szarkoplazmába az extracelluláris folyadékból, és lassan távoznak is, miután az idegimpulzusok már nem érkeznek. Ezért a simaizomrostok lassan összehúzódnak és lassan ellazulnak.
A csontváz általános áttekintése emberi izmok. A törzs izmai(2.6. és 2.7. ábra) tartalmazzák a mellkas, a hát és a has izmait. A mellkas izmai részt vesznek a felső végtagok mozgásában, emellett akaratlagos és akaratlan légzőmozgásokat is biztosítanak. A mellkas légzőizmoit külső és belső bordaközi izmoknak nevezzük. A légzőizmok közé tartozik a rekeszizom is. A hátizmok felületes és mély izmokból állnak. A felületesek bizonyos mozgásokat biztosítanak a felső végtagokon, a fejen és a nyakon. A mélyek („törzs egyenirányítói”) a csigolyák tövisnyúlványaihoz kapcsolódnak, és a gerinc mentén húzódnak. A hátizmok részt vesznek a test függőleges helyzetének megőrzésében, erős feszültséggel (összehúzódással) a testet hátrahajlítják. A hasizmok fenntartják a nyomást a hasüregben (hasi), részt vesznek bizonyos testmozgásokban (törzs előrehajlítása, hajlítás és oldalra fordulás), valamint a légzési folyamat során.
A fej és a nyak izmai - a fej és a nyak utánzása, rágása és mozgatása. Az arcizmok egyik végén a csonthoz, a másikon az arc bőréhez csatlakoznak, egyesek a bőrben kezdődhetnek és végződhetnek. Az arcizmok biztosítják az arcbőr mozgását, tükrözik az ember különféle mentális állapotait, kísérik a beszédet és fontosak a kommunikációban. Amikor a rágóizmok összehúzódnak, az alsó állkapocs előre és oldalra mozog. A nyakizmok részt vesznek a fej mozgásában. A hátsó izomcsoport, beleértve a fej hátsó izmait is, tónusos (a „tónus” szóból) összehúzódással függőleges helyzetben tartja a fejet.

Rizs. 2.6. A test elülső felének izmai (Sylvanovich szerint):
1 - temporális izom, 2 - rágóizom, 3 - sternocleidomastoideus izom, 4 - nagy mellizom, 5 - középső pikkely izom, b - külső ferde hasizom, 7 - vastus medialis, 8 - vastus lateralis, 9 - rectus femoris izom, 10 - Sartorius, 11 - érzékeny izom 12 - belső ferde hasizom, 13 - rectus hasizom, 14 - bicepsz brachii izom, 15 ~ külső bordaközi izmok, 16 - orbicularis oris izom, 17 - orbicularis oculi izom, 18 - frontalis izom
A felső végtagok izmai biztosítsa a vállöv, a váll, az alkar mozgását és mozgassa a kezet és az ujjakat. A fő antagonista izmok a váll bicepsz (hajlító) és tricepsz (extensor) izmai. A felső végtag és mindenekelőtt a kéz mozgása rendkívül változatos. Ez annak a ténynek köszönhető, hogy a kéz emberi munkaszervként szolgál.

Rizs. 2.7. A test hátsó felének izmai (Sylvanovich szerint):
1 - rombusz izom, 2 - egyenirányító törzs, 3 - a gluteális izomzat mély izmai, 4 - bicepsz femoris izom, 5 - vádli izom, 6 - Achilles-ín, 7 - gluteus maximus izom, 8 - latissimus skipae izom, 9 - deltoid, 10 - trapéz izom
Az alsó végtagok izmai biztosítják a csípő, a lábszár és a lábfej mozgását. A combizmok fontos szerepet játszanak az egyenes testhelyzet megőrzésében, de az embernél fejlettebbek, mint más gerinceseknél. Az alsó lábszár mozgását végző izmok a combon helyezkednek el (például a négyfejű izom, amelynek funkciója az alsó láb térdízületben történő meghosszabbítása; ennek az izomnak az antagonistája a biceps femoris izom). A lábfejet és a lábujjakat az alsó lábszárban és a lábfejben található izmok hajtják. A lábujjak hajlítását a talpon található izmok összehúzódása, a láb és a lábfej elülső felületének izomzata nyújtja. A comb, a láb és a lábfej számos izma részt vesz az emberi test függőleges helyzetben tartásában.
VAL VELizomrostok szerkezete és összehúzódása.
Az izomösszehúzódás egy élő rendszerben mechanokémiai folyamat. A modern tudomány a biológiai mobilitás legtökéletesebb formájának tartja. A biológiai objektumok „kifejlesztették” az izomrostok összehúzódását, mint a térben való mozgás egyik módját (ami jelentősen kibővítette életképességüket).
Az izomösszehúzódást egy feszülési fázis előzi meg, amely a kémiai energia közvetlen és jó hatásfokkal (30-50%) mechanikai energiává alakításával végzett munka eredménye. A potenciális energia felhalmozódása a feszülési fázisban az izomzatot a lehetséges, de még nem realizált összehúzódás állapotába hozza.
Az állatok és az emberek (és az emberek azt hiszik, hogy már alaposan tanulmányozták) két fő izomtípus: csíkozott és sima. Harántcsíkolt izmok vagy csontváz a csontokhoz kötődik (kivéve a szívizom harántcsíkolt rostjait, amelyek összetételükben különböznek a vázizmoktól). Sima izmok támogatja a belső szervek és a bőr szöveteit, valamint kialakítja az erek falának izmait, valamint a beleket.
A sport biokémiájában tanulnak vázizmok, „kifejezetten felelős” a sporteredményekért.
Egy izom (mint egy makroobjektumhoz tartozó makroképződmény) egyedekből áll izomrostok(mikroképződmények). Több ezer van belőlük egy izomban, ennek megfelelően az izomerőfeszítés olyan szerves érték, amely számos egyedi rost összehúzódását összegzi. Háromféle izomrost létezik: fehér gyors rángatózás , közbülsőÉs piros lassú rándulás. A rostok típusai különböznek az energiaellátásuk mechanizmusában, és különböző motoros neuronok irányítják őket. Az izomtípusok a rosttípusok arányában különböznek.
Külön izomrost - fonalszerű acelluláris képződmény - egyszerű. A szimplaszt „nem úgy néz ki, mint egy sejt”: erősen megnyúlt alakja van, hossza 0,1-2-3 cm, a Sartorius izomzata legfeljebb 12 cm, vastagsága 0,01-0,2 mm. A szimplasztot egy héj veszi körül - sarcolemma, melynek felületéhez több mozgatóideg vége közeledik. A Sarcolemma egy kétrétegű lipoprotein membrán (10 nm vastag), amelyet kollagénrostok hálózata erősít meg. Amikor összehúzódás után ellazulnak, visszaállítják a szimplaszt eredeti alakját (4. ábra).
Rizs. 4. Egyedi izomrost.
A szarkolemma-membrán külső felületén az elektromos membránpotenciál mindig megmarad, nyugalmi állapotban is 90-100 mV. A potenciál jelenléte szükséges feltétele az izomrostok szabályozásának (mint az autó akkumulátora). A potenciál az anyagok aktív (értsd energiafelhasználás - ATP) membránon keresztüli átvitele és szelektív permeabilitása miatt jön létre (akit akarok, azt beengedem vagy kiengedem) elv szerint. ). Ezért a szimplaszt belsejében egyes ionok és molekulák nagyobb koncentrációban halmozódnak fel, mint azon kívül.
A szarkolemma jól átereszti a K + ionokat - belül halmozódnak fel, és a Na + ionok kívülről távoznak. Ennek megfelelően a Na + ionok koncentrációja az intercelluláris folyadékban nagyobb, mint a szimplaszton belüli K + ionok koncentrációja. A pH-eltolódás a savas oldalra (például a tejsav képződése során) növeli a szarkolemma permeabilitását a nagy molekulatömegű anyagok (zsírsavak, fehérjék, poliszacharidok) számára, amelyek normál esetben nem jutnak át rajta. Az alacsony molekulatömegű anyagok (glükóz, tej- és piroszőlősav, ketontestek, aminosavak, rövid peptidek) könnyen átjutnak (diffundálnak) a membránon.
A simplast belső tartalma – szarkoplazma– Ez egy kolloid fehérjeszerkezet (az állaga zselére emlékeztet). Szuszpendált állapotban glikogén zárványokat, zsírcseppeket tartalmaz, és különféle szubcelluláris részecskék „beépültek”: magok, mitokondriumok, myofibrillumok, riboszómák és mások.
Összehúzódó „mechanizmus” a szimplaszton belül – myofibrillumok. Ezek vékony (Ø 1-2 mikron) izomszálak, hosszúak - majdnem megegyeznek az izomrost hosszával. Megállapítást nyert, hogy az edzetlen izmok szimplasztjaiban a myofibrillumok nem rendezetten, a szimplaszt mentén helyezkednek el, hanem szórással és eltérésekkel, az edzetteknél pedig a hossztengely mentén orientálódnak és szintén csoportosulnak. kötegek, mint a kötelekben. (A mesterséges és szintetikus szálak fonásánál a polimer makromolekulái kezdetben nem szigorúan a szál mentén helyezkednek el, és a sportolókhoz hasonlóan ismételt visszatekeréssel „kitartóan edződnek” - helyesen orientálva - a szálak tengelye mentén: lásd a hosszú műhelyek a ZIV-ben és Khimvoloknóban).
Fénymikroszkóp alatt megfigyelhető, hogy a myofibrillák valóban „csíkozottak”. Felváltva világos és sötét területeket - korongokat. Sötét felnik A Az (anizotrop) fehérjék többet tartalmaznak, mint a könnyű korongok én (izotróp). Membránokkal keresztezett fénykorongok Z (telofragmák) és a kettő közötti myofibrillum egy szakasza Z - membránoknak nevezik sarcomere. A myofibrill 1000-1200 szarkomerből áll (5. ábra).
Az izomrost összehúzódása összességében egyedi összehúzódásokból áll szarkomerek. Mindegyiket külön-külön összehúzva a szarkomerek együtt egy integrált erőt hoznak létre, és mechanikus munkát végeznek az izom összehúzására.
A szarkomer hossza nyugalmi állapotban 1,8 µm és mérsékelt 1,5 µm, teljes összehúzódás esetén pedig 1 µm között változik. A szarkomerek sötét és világos korongjai protofibrillumot (miofilamentumot) tartalmaznak - fehérjeszál-szerű struktúrákat. Két típusban találhatók: vastag (Ø – 11 – 14 nm, hosszúság – 1500 nm) és vékony (Ø – 4 – 6 nm, hosszúság – 1000 nm).

Rizs. 5. Myofibrill terület.
Könnyű kerekek ( én ) csak vékony protofibrillákból és sötét korongokból állnak ( A ) – kétféle protofibrillumból: vékony, membránnal összeerősített és vastag, külön zónában koncentrált ( H ).
Amikor a szarkomér összehúzódik, a sötét korong hossza ( A ) nem változik, és a fénykorong hossza ( én ) csökken, ahogy a vékony protofibrillumok (világos korongok) a vastagok (sötét korongok) közötti térbe kerülnek. A protofibrillumok felületén speciális kinövések - adhéziók (kb. 3 nm vastag) vannak. A „munkahelyzetben” kapcsolatot (kereszthidakat) képeznek a protofibrillák vastag és vékony szálai között (6. ábra). Szerződéskötéskor Z - a membránok a vastag protofibrillumok végein támaszkodnak, a vékony protofibrillumok pedig még a vastagok köré is tekerhetnek. A szuperkontrakció során a szarkomer közepén lévő vékony filamentumok végei felkunkorodnak, a vastag protofibrillumok végei pedig összetörnek.

Rizs. 6. Adhéziók kialakulása aktin és miozin között.
Az izomrostok energiaellátása segítségével történik szarkoplazmatikus retikulum(más néven - szarkoplazmatikus retikulum) – hosszanti és keresztirányú csövek, membránok, buborékok, rekeszek rendszerei.
A szarkoplazmatikus retikulumban különböző biokémiai folyamatok szervezetten és kontrolláltan mennek végbe, a hálózat mindent együtt és minden myofibrillumot külön-külön lefed. A retikulum riboszómákat tartalmaz, fehérjék szintézisét végzik, és mitokondriumok - „sejtenergia-állomások” (az iskolai tankönyvben meghatározottak szerint). Tulajdonképpen mitokondriumok miofibrillumok közé ágyazva, ami optimális feltételeket teremt az izomösszehúzódási folyamat energiaellátásához. Megállapították, hogy az edzett izmokban a mitokondriumok száma nagyobb, mint az azonos edzetlen izmokban.
Az izmok kémiai összetétele.
Vízzel elhagyja az izomtömeg 70-80%-át.
Mókusok. A fehérjék az izomtömeg 17-21%-át teszik ki: az összes izomfehérje hozzávetőlegesen 40%-a a myofibrillumokban, 30%-a a szarkoplazmában, 14%-a a mitokondriumokban, 15%-a a szarkolemmában, a többi a sejtmagokban és más sejtszervecskékben koncentrálódik.
Az izomszövet enzimeket tartalmaz miogén fehérjék csoportok, myoalbumin– tartalék fehérje (tartalma az életkorral fokozatosan csökken), vörös fehérje mioglobin- kromoprotein (ezt izom hemoglobinnak hívják, több oxigént köt meg, mint a vér hemoglobinja), valamint globulinok, myofibrilláris fehérjék. A myofibrilláris fehérjék több mint fele miozin, körülbelül egynegyede - aktin, a többi tropomiozin, troponin, α- és β-aktinin, enzimek kreatin-foszfokináz, deamináz és mások. Az izomszövet tartalmaz nukleárismókusok- nukleoproteinek, mitokondriális fehérjék. A fehérjékben stroma,összefonódó izomszövet - a fő rész - kollagénÉs elasztin sarcolemma, valamint myostrominok (kapcsolódóan Z -membránok).
Ban benelőoldható nitrogénvegyületek. Az emberi vázizmok különféle vízben oldódó nitrogénvegyületeket tartalmaznak: ATP, 0,25-0,4% kreatin-foszfát (CrP)– 0,4-ről 1%-ra (edzéssel nő a mennyisége), bomlástermékeik ADP, AMP, kreatin. Ezenkívül az izmok tartalmaznak egy dipeptidet is karnozin, körülbelül 0,1-0,3%, részt vesz az izomteljesítmény helyreállításában fáradtság esetén; karnitin, felelős a zsírsavak sejtmembránokon történő szállításáért; aminosavak, és köztük a glutamin dominál (ez magyarázza-e a mononátrium-glutamát használatát, olvassa el az ízesítők összetételét, hogy az ételnek a hús ízét adja); purinbázisok, karbamid és ammónia. A vázizom is körülbelül 1,5% foszfatidok, amelyek részt vesznek a szöveti légzésben.
Nitrogénmentes kapcsolatokat. Az izmok szénhidrátokat, glikogént és anyagcseretermékeit, valamint zsírokat, koleszterint, ketontesteket és ásványi sókat tartalmaznak. Az étrendtől és az edzés mértékétől függően a glikogén mennyisége 0,2-3% között változik, míg az edzés növeli a szabad glikogén tömegét. Az állóképességi edzés során a raktározó zsírok felhalmozódnak az izmokban. A fehérjékhez kötött zsír körülbelül 1%-ot tesz ki, az izomrostok membránja pedig akár 0,2%-ot is tartalmazhat.
Ásványok. Az izomszövetben található ásványi anyagok az izomtömeg körülbelül 1-1,5%-át teszik ki; ezek főként kálium-, nátrium-, kalcium- és magnéziumsók. Az ásványi ionok, mint a K +, Na +, Mg 2+, Ca 2+, Cl -, HP0 4 ~ létfontosságú szerepet játszanak az izomösszehúzódás során zajló biokémiai folyamatokban (a „sport” kiegészítőkben és az ásványvízben szerepelnek).
Az izomfehérjék biokémiája.
Az izmok fő kontraktilis fehérje az miozin A kifejezés fibrilláris fehérjékre vonatkozik (molekulatömege körülbelül 470 000). A miozin fontos tulajdonsága, hogy komplexeket képez az ATP- és ADP-molekulákkal (amely lehetővé teszi az energia „elvételét” az ATP-ből), valamint az aktin fehérjével (ami lehetővé teszi az összehúzódás fenntartását).
A miozin molekula negatív töltésű, és specifikusan kölcsönhatásba lép a Ca ++ és Mg ++ ionokkal. A miozin Ca++ ionok jelenlétében felgyorsítja az ATP hidrolízisét, így enzimatikus adenozin-trifoszfát aktivitás:
miozin-ATP+H2O → miozin + ADP + H3PO4 + munka(energia 40 kJ/mol)
A miozin fehérjét két azonos, hosszú polipeptid α-lánc alkotja, amelyek kettős hélixszerűen csavarodnak össze, 7. ábra. A proteolitikus enzimek hatására a miozin molekula két részre szakad. Egyik része adhézión keresztül képes kötődni az aktinhoz, aktomiozint képezve. Ez a rész felelős az adenozin-trifoszfatáz aktivitásért, ami a környezet pH-jától függ, az optimum pH 6,0 - 9,5, valamint a KCl koncentrációja. Az aktomiozin komplex ATP jelenlétében szétesik, de szabad ATP hiányában stabil. A miozinmolekula második része szintén két csavart hélixből áll, amelyek elektrosztatikus töltés hatására protofibrillumokká kötik a miozin molekulákat.

Rizs. 7. Az aktomiozin szerkezete.
A második legfontosabb kontraktilis fehérje az aktin(7. ábra). Három formában létezhet: monomer (globuláris), dimer (globuláris) és polimer (fibrilláris). A monomer globuláris aktin, amikor polipeptidláncai szorosan tömör gömb alakú szerkezetbe tömörülnek, az ATP-vel asszociálódnak. Az ATP felhasításával az aktin monomerek - A dimereket képeznek, beleértve az ADP-t: A - ADP - A. A polimer fibrilláris aktin egy dimerekből álló kettős hélix, ábra. 7.
A globuláris aktin K + és Mg ++ ionok jelenlétében fibrilláris aktinná alakul, az élő izmokban pedig a fibrilláris aktin dominál.
A myofibrillumok jelentős mennyiségű fehérjét tartalmaznak tropomiozin, amely két α-helikális polipeptid láncból áll. Nyugvó izmokban komplexet képez az aktinnal és blokkolja annak aktív központjait, mivel az aktin képes kötődni Ca ++ ionokhoz, amelyek megszüntetik ezt a blokádot.
Molekuláris szinten a szarkomer vastag és vékony protofibrillumai elektrosztatikusan kölcsönhatásba lépnek, mivel speciális területeik vannak - kinövések és kiemelkedések -, ahol töltés képződik. Az A-korong régióban vastag protofibrillumok hosszirányban orientált miozinmolekulák kötegéből épülnek fel, vékony protofibrillumok sugárirányban helyezkednek el a vastagok körül, így a többszálú kábelhez hasonló szerkezetet alkotnak. A vastag protofibrillumok központi M-sávjában a miozinmolekulákat „farkuk” köti össze, kiálló „fejük” - a kinövések különböző irányokba irányulnak, és szabályos spirálvonalak mentén helyezkednek el. Valójában velük szemben a fibrilláris aktinspirálokban, egymástól bizonyos távolságra, monomer aktingömbök is kiállnak. Mindegyik kiemelkedésnek van aktív központ, ami miatt lehetséges a miozinnal való adhézió kialakulása. A szarkomerek Z-membránjai (mint a váltakozó talapzatok) vékony protofibrillumot tartanak össze.
Az összehúzódás és relaxáció biokémiája.
Az izomban az összehúzódás során fellépő ciklikus biokémiai reakciók biztosítják a „fejek” – a vastag protofibrillumok miozinmolekuláinak kinövései és a vékony protofibrillumok aktív központjai – közötti kiemelkedések ismételt kialakulását és pusztulását. Az adhézióképzés és az aktin filamentum miozin filamentum mentén történő mozgatása pontos szabályozást és jelentős energiafelhasználást igényel. A valóságban a rostok összehúzódásának pillanatában percenként körülbelül 300 adhézió képződik minden aktív központban - kiemelkedésben.
Amint azt korábban megjegyeztük, csak az ATP-energiát lehet közvetlenül az izomösszehúzódás mechanikai munkájává alakítani. A miozin enzimatikus központja által hidrolizált ATP komplexet képez a teljes miozin fehérjével. Az ATP-miozin komplexben az energiával telített miozin megváltoztatja szerkezetét, és ezzel a külső „dimenziókat”, és ezáltal mechanikai munkát végez a miozin filamentum növekedésének lerövidítése érdekében.
Nyugalmi izomban a miozin továbbra is kötődik az ATP-hez, de Mg++ ionokon keresztül az ATP hidrolitikus hasítása nélkül. A miozin és az aktin közötti adhézió kialakulását nyugalmi állapotban megakadályozza a tropomiozin troponinnal alkotott komplexe, amely blokkolja az aktin aktív központjait. A blokád fennmarad, és az ATP nem bomlik le, miközben a Ca++ ionok megkötődnek. Amikor idegimpulzus érkezik egy izomrosthoz, az felszabadul impulzus adó- neurohormon acetilkolin. A Na+ ionok semlegesítik a szarkolemma belső felületén lévő negatív töltést és depolarizálják azt. Ebben az esetben Ca++ ionok szabadulnak fel és kötődnek a troponinhoz. A troponin viszont elveszíti töltését, aminek következtében az aktív központok - az aktin filamentumok kiemelkedései - felszabadulnak, és az aktin és a miozin között adhéziók keletkeznek (mivel a vékony és vastag protofibrillumok elektrosztatikus taszítása már megszűnt). Most, Ca ++ jelenlétében, az ATP kölcsönhatásba lép a miozin enzimatikus aktivitásának központjával és hasad, és a transzformáló komplex energiáját használják fel az adhézió csökkentésére. A molekuláris események fent leírt láncolata hasonló a mikrokondenzátort feltöltő elektromos áramhoz, elektromos energiája azonnal mechanikai munkává alakul a helyszínen, és újra kell tölteni (ha tovább akarunk lépni).
A ragasztóanyag felszakadása után az ATP nem hasad, hanem ismét enzim-szubsztrát komplexet képez a miozinnal:
M–A + ATP -----> M – ATP + A vagy
M–ADP–A + ATP ----> M–ATP + A + ADP
Ha ebben a pillanatban új idegimpulzus érkezik, akkor a „feltöltési” reakciók ismétlődnek, ha a következő impulzus nem érkezik meg, az izom ellazul. Az összehúzott izom relaxáció után az eredeti állapotba való visszatérését az izomsztrómában lévő fehérjék rugalmas erői biztosítják. Az izomösszehúzódás modern hipotéziseit felállítva a tudósok azt sugallják, hogy az összehúzódás pillanatában az aktin filamentumok a miozin filamentumok mentén csúsznak, és ezek megrövidülése a kontraktilis fehérjék térszerkezetének megváltozása (a hélix alakjának változása) miatt is lehetséges.
Nyugalomban az ATP lágyító hatású: miozinnal kombinálva megakadályozza annak aktinnal való adhézióinak kialakulását. Az izomösszehúzódás során lebomolva az ATP energiát biztosít az adhéziók lerövidítésének folyamatához, valamint a „kalciumpumpa” munkájához - a Ca ++ ionok ellátásához. Az ATP lebomlása az izomban nagyon nagy sebességgel megy végbe: percenként akár 10 mikromol/1 g izom is. Mivel az izomzat összes ATP-tartaléka kicsi (lehet, hogy maximum 0,5-1 mp-es munkára elegendő), a normál izomműködés biztosításához az ATP-t ugyanolyan ütemben kell helyreállítani, mint ahogyan lebomlik.