Единици на живите същества: хлоропласти. Хлоропластите, тяхната структура, химичен състав и функции 1 структура и функции на хлоропластите
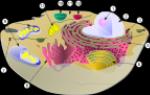
(Гръцки "хлорос" - зелен) - двойномембранни органели с доста сложна структура, съдържащи хлорофил и извършващи фотосинтеза. Характерен само за растителните клетки (фиг. 1). Във водораслите носителите на хлорофил са хроматофорите - предшествениците на пластидите; те се срещат и в животното - зелена еуглена (различни форми). Хлоропластите на висшите растения имат формата на двойноизпъкнала леща, която най-ефективно улавя светлината. В една клетка има средно 10-30 (до 1000) хлоропласта. Дължината на пластида е 5-10 микрона, дебелина - 1-3, ширина - 2-4 микрона. Хлоропластите са покрити с външна гладка мембрана, докато вътрешната мембрана образува структури, наречени тилакоиди (торбички) в кухината на пластида. Дисковидните тилакоиди образуват грана, а тръбовидните тилакоиди образуват строма тилакоиди, свързващи всички грани в една система. Една грана съдържа от няколко до 50 тилакоиди, а броят на граните в хлоропласта достига 40-60. Пространството между стромалните тилакоиди и граната е изпълнено с „основното вещество“ - стромата. състоящ се от протеини, липиди, въглехидрати, ензими, АТФ. В допълнение, стромата съдържа пластидна ДНК. РНК, рибозоми. Тилакоидните мембрани имат типична структура, но за разлика от други органели съдържат оцветяващи вещества - хлорофилни (зелени) пигменти и каротеноиди (червено-оранжево-жълти). Хлорофил- основният пигмент, свързан с глобуларни протеини в протеиново-пигментни комплекси, разположени от външната страна на тилакоидната мембрана на грана. Каротеноиди- допълнителни пигменти се намират в липидния слой на мембраната, където не се виждат, тъй като са разтворени в мазнини. Но тяхното местоположение точно съответства на протеиново-пигментния комплекс, така че пигментите в мембраните не образуват непрекъснат слой, а са разпределени мозаечно. Структурата на хлоропластите е тясно свързана с тяхната функция. В тях протича фотосинтеза; Светлинните реакции протичат върху зърнестите тилакоидни мембрани, а въглеродната фиксация се извършва в стромата (тъмни реакции). Хлоропласти- полуавтономни органели, в които се синтезират собствени протеини, но те не могат да живеят дълго време извън клетката, тъй като са под общия контрол на клетъчното ядро. Те се възпроизвеждат чрез делене на половина или могат да се образуват от пропластиди или левкопласти. Пропластидите се предават през зиготата под формата на много малки тела, диаметърът им е 0,4-1,0 микрона, те са безцветни и покрити с двойна мембрана. Пропластидите се намират в клетките на растежния конус на стъблото и корена и в примордиите на листата. В зелените органи - листа, стъбла - те се превръщат в хлоропласти. В края на жизнения цикъл хлорофилът се унищожава (обикновено при промяна на дневната светлина и понижаване на температурата), някои от хлоропластите се превръщат в хромопласти - зелените листа и плодове стават червени или жълти и след това падат.
Ориз. 1. Структура:a - хлоропласт, b - левкопласт, c - хромопласт; 1 - външна мембрана, 2 - вътрешна мембрана, 3 - метрика (строма), 4 - стромални тилакоиди (ламели), 5 - грана, c - тилакоидна грана, 7 - нишестено зърно, 8 - каротеноиди в липидни капчици, 9 - ДНК, 10 - рибозоми, 11 - срутващи се мембранни структури
фотосинтезата се извършва в специализирани клетъчни органели - хлоропласти. Хлоропластите на висшите растения имат двойноизпъкнала форма лещи(диск), който е най-удобен за абсорбиране на слънчева светлина. Техният размер, количество и местоположение напълно отговарят на предназначението им: максимално ефективно усвояване на слънчевата енергия и максимално пълно усвояване на въглерода. Установено е, че броят на хлоропластите в една клетка се измерва в десетки. Това осигурява високо съдържание на тези органели на единица листна повърхност. Да, на 1 мм 2 листа от боб отчетени 283 хилядихлоропласти, в слънчогледа - 465 хиляди. Диаметърхлоропласти средно 0,5-2 микрона.
Структура на хлоропластамного сложно. Подобно на ядрото и митохондриите, хлоропластът е заобиколен от обвивка, състояща се от две липопротеинови мембрани. Вътрешната среда е представена от относително хомогенна субстанция - матрица или строма , който е проникнат от мембрани - ламели (ориз.). Ламелите, свързани една с друга, образуват мехурчета - тилакоиди . Плътно прилепнали един към друг, се образуват тилакоиди зърна , които се различават дори под светлинен микроскоп. На свой ред граните на едно или няколко места са обединени помежду си с помощта на междугранални нишки - стромални тилакоиди.
Свойства на хлоропластите: способни да променят ориентацията си и се движи. Например, под въздействието на ярка светлина, хлоропластите обръщат тясната страна на диска към падащите лъчи и се придвижват към страничните стени на клетките. Хлоропластите се движат към по-високи концентрации на CO 2 в клетката. През деня обикновено се нареждат покрай стените, а през нощта потъват на дъното на клетката.
Химичен съставхлоропласти: вода - 75%; 75-80% от общото количество сухо вещество е орг. съединения, 20-25% минерал.
Структурната основа на хлоропластите е катерици (50-55 % суха маса), половината от тях са водоразтворими протеини. Такова високо съдържание на протеин се обяснява с техните разнообразни функции в хлоропластите (структурни мембранни протеини, ензимни протеини, транспортни протеини, контрактилни протеини, рецепторни протеини).
Най-важните компоненти на хлоропластите са липиди , (30-40% суха м.). Хлоропластните липиди са представени от три групи съединения.
Структурни компоненти на мембраните, които са представени от амфипатични липоиди и се характеризират с високо съдържание (повече от 50%) на галактолипиди и сулфолипиди. Характеризира се фосфолипидният състав липса на фосфатидилетаноламини високо съдържание фосфатидилглицерол(повече от 20%). Над 60 % съставът на течните кристали отчита линоловакиселина.
Фотосинтетични пигментихлоропласти - хидрофобни вещества, свързани с липоиди(водоразтворими пигменти в клетъчния сок). Висшите растения съдържат 2 форми зеленопигменти: хлорофил аИ хлорофилbи 2 форми на жълти пигменти: каротиниИ ксантофили(каротеноиди). Хлорофилът играе роля фотосенсибилизатори, други пигменти разширяват спектъра на фотосинтезата поради по-пълното усвояване на PAR. Каротеноидите предпазват хлорофила от фотоокисление, участват в транспорт на водород, образуван при фотолизата на водата.
Мастноразтворими витамини - ергостерол(провитамин D), вит д, ДА СЕ- концентрирани почти изцяло в хлоропластите, където участват в превръщането на светлинната енергия в химична. Цитозолът на листните клетки съдържа главно водоразтворими витамини. Така в спанака съдържанието на аскорбинова киселина в хлоропластите е 4-5 пъти по-малко, отколкото в листата.
Хлоропластите на листата съдържат значително количество РНК и ДНК . NC съставляват приблизително 1% от сухото тегло на хлоропластите (РНК - 0,75%, ДНК - 0,01-0,02%). Геномът на хлоропласта е представен от кръгова ДНК молекула с дължина 40 µm с молекулно тегло 108, кодираща 100-150 средно големи протеини. Хлоропластните рибозоми съставляват 20 до 50% от общата популация рибозоми в клетката. По този начин хлоропластите имат своя собствена система за синтез на протеини. За нормалното функциониране на хлоропластите обаче е необходимо взаимодействие между ядрения и хлоропластния геном. Ключовият ензим на фотосинтезата, RDP карбоксилазата, се синтезира под двоен контрол - ДНК на ядрото и хлоропласта.
Въглехидратине са конституционни вещества на хлоропласта. Те са представени от фосфорни естери на захари и продукти на фотосинтезата. Следователно съдържанието на въглехидрати в хлоропластите варира значително (от 5 до 50%). В активно функциониращите хлоропласти въглехидратите обикновено не се натрупват, възниква бързото им изтичане. С намаляване на нуждата от фотосинтетични продукти в хлоропластите се образуват големи нишестени зърна. В този случай съдържанието на нишесте може да се увеличи до 50 % сухата маса и активността на хлоропластите ще намалеят.
Минерали. Самите хлоропласти съставляват 25-30% от листната маса, но съдържат до 80 % Fe, 70-72 - MgИZn, 50 - Cu, 60 % оксъдържащи се в листните тъкани. Това се обяснява с високата и разнообразна ензимна активност на хлоропластите (включително простетични групи и кофактори). Mgе част от хлорофила. окстабилизира мембранните структури на хлоропластите.
Появата и развитието на хлоропластите . Хлоропластите се образуват в меристемни клетки от първоначални частици или рудиментарни пластиди (фиг.). Първоначалната частица се състои от амебоиден поток, заобиколен от двумембранна обвивка. С нарастването на клетката първоначалните частици се увеличават по размер и придобиват формата на двойноизпъкнала леща, а в стремчетата се появяват малки нишестени зърна. В същото време вътрешната мембрана започва да расте, образувайки гънки (инвагинации), от които излизат везикули и тръби. Такива образувания се наричат пропластиди . За по-нататъшното им развитие е необходима светлина. На тъмно се образуват етиопласти , в който се образува мембранна решетъчна структура - проламеларното тяло. На светлина се образуват вътрешните мембрани на пропластидите и етиопластите режеща система. В същото време новообразуваните молекули на хлорофила и други пигменти също се вграждат в граната на светлина. Така структури, които са подготвени да функционират на светлина, се появяват и развиват само в нейно присъствие.
Наред с хлоропластите има редица други пластиди, които се образуват или директно от пропластиди, или един от друг чрез взаимни трансформации ( ориз.). Те включват амилопласти, натрупващи нишесте ( левкопласти) И хромопластисъдържащи каротеноиди. В цветята и плодовете хромопластите възникват рано в развитието на пропластидите. Хромопластите на есенната зеленина са продукти от разграждането на хлоропластите, в които пластоглобулите действат като каротноидни носещи структури.
Пигменти хлоропласти, участващи в улавянето на светлинна енергия, както и ензими, необходими за светлинната фаза фотосинтеза, вградена в мембранитилакоиди.
Ензими , които катализират многобройни реакции на редукционния цикъл на въглехидратите (темпова фаза на фотосинтезата), както и различни биосинтези, включително биосинтезата на протеини, липиди, нишесте, присъстват главно в стромата, някои от тях са периферни ламелни протеини.
Структурата на зрелите хлоропласти е еднаква във всички висши растения, както и в клетките на различни органи на едно и също растение (листа, зелени корени, кора, плодове). В зависимост от функционалното натоварване на клетките, физиологичното състояние на хлоропластите и тяхната възраст се разграничава степента на тяхната вътрешна структура: размер, брой зърна, връзка между тях. И така, в заключение устични клеткиосновната функция на хлоропластите е фоторегулацияустични движения. Хлоропластите нямат строга гранулирана структура, те съдържат големи нишестени зърна, подути тилакоиди и липофилни глобули. Всичко това показва ниското им енергийно натоварване (тази функция се изпълнява от митохондриите). Друга картина се наблюдава при изследване на хлоропластите на плодовете на зелените домати. Наличност добре развита гранулирана системапоказва високо функционално натоварване на тези органели и вероятно значителен принос на фотосинтезата по време на образуването на плода.
Промени, свързани с възрастта: Младите се характеризират с ламеларна структура; в това състояние хлоропластите могат да се възпроизвеждат чрез делене. При зрелите грановата система е добре изразена. При стареещите индивиди стромалните тилакоиди се разкъсват, връзката между грана намалява и впоследствие се наблюдава разпадане на хлорофила и разрушаване на грана. В есенната зеленина разграждането на хлоропластите води до образуването хромопласти .
Структура на хлоропласта лабилен и динамичен , той отразява всички условия на живот на растението. Голямо влияние оказва режимът на минерално хранене на растенията. Ако има недостиг нхлоропластите стават 1,5-2 пъти по-малки, недостиг ПИ Снарушава нормалната структура на ламели и грани, едновременен дефицит нИ окводи до препълване на хлоропластите с нишесте поради нарушаване на нормалното изтичане на асимилати. Ако има недостиг окструктурата на външната мембрана на хлоропласта е нарушена. За поддържане на структурата на хлоропласта е необходима и светлина; на тъмно зърнестите и стремалните тилакоиди постепенно се разрушават.
Те са групирани в grana, които са купчини от дисковидни тилакоиди, сплескани и плътно притиснати един към друг. Граните са свързани с помощта на ламели. Пространството между мембраната на хлоропласта и тилакоидите се нарича строма. Стромата съдържа хлоропластни молекули РНК, пластидна ДНК, рибозоми , нишестеназърна и ензими Цикъл на Калвин.
Произход
Произходът на хлоропластите чрез симбиогенеза вече е общоприет. Смята се, че хлоропластите възникнаот цианобактерии, тъй като те са двумембранна органела, имат собствена затворена кръгова ДНК и РНК, пълноценен апарат за синтез на протеини (и рибозоми от прокариотен тип - 70S), размножават бинарно делене, а тилакоидните мембрани са подобни на мембраните на прокариотите (чрез наличието на киселинни липиди) и приличат на съответните органели в цианобактериите. U глаукофитиводорасли, вместо типични хлоропласти, клетките съдържат цианела- цианобактерии, които в резултат на ендосимбиоза са загубили способността си да съществуват независимо, но са запазили частично цианобактериалната клетъчна стена.
Продължителността на това събитие се оценява на 1 - 1,5 милиарда години.
Някои групи организми са получили хлоропласти в резултат на ендосимбиоза не с прокариотни клетки, а с други еукариоти, които вече са имали хлоропласти. Това обяснява наличието на повече от две мембрани в мембраната на хлоропласта на някои организми. Най-вътрешната от тези мембрани се тълкува като изгубена клетъчна стенаобвивката на цианобактерия, външната е като стената на симбионтофорната вакуола на гостоприемника. Междинните мембрани принадлежат на редуциран еукариотен организъм, който е влязъл в симбиоза. В някои групи в перипластидното пространство между втората и третата мембрана има нуклеоморф, силно редуцирано еукариотно ядро.
Хлоропластен модел
Структура
В различните групи организми хлоропластите се различават значително по размер, структура и брой в клетката. Структурните характеристики на хлоропластите са големи таксономичензначение .
Хлоропластна обвивка
В различните групи организми хлоропластната мембрана се различава по структура.
При глаукоцистофитите, червените и зелените водорасли и при висшите растения черупката се състои от две мембрани. При други еукариотни водорасли хлоропластът е допълнително заобиколен от една или две мембрани. При водораслите, които имат четиримембранни хлоропласти, външната мембрана обикновено се слива с външната мембрана на ядрото.
Перипластидно пространство
Ламела и тилакоиди
Ламелите свързват тилакоидните кухини
Пиреноиди
Пиреноидите са центрове на синтеза на полизахариди в хлоропластите. Структурата на пиреноидите е разнообразна и те не винаги са морфологично изразени. Те могат да бъдат интрапластидни или подобни на стебла, изпъкнали в цитоплазмата. В зелените водорасли и растенията пиреноидите се намират вътре в хлоропласта, което е свързано с вътрепластидното съхранение на нишесте.
Стигма
Стигми или оцели се намират в хлоропластите на клетките на подвижните водорасли. Разположен близо до основата на флагела. Стигмите съдържат каротеноидии могат да работят като фоторецептори.
Вижте също
Бележки
Коментари
Бележки
Литература
- Белякова Г. А.Водорасли и гъби // Ботаника: в 4 тома / Белякова Г. А., Дяков Ю. Т., Тарасов К. Л. - М.: Издателски център "Академия", 2006. - Т. 1. - 320 с. – 3000 бр. - ISBN 5-7695-2731-5
- Карпов С.А.Структурата на протистната клетка. - Санкт Петербург. : ТЕСА, 2001. - 384 с. - 1000 бр. - ISBN 5-94086-010-9
- Лий, Р. Е.Фикология, 4-то издание. - Cambridge: Cambridge University Press, 2008. - 547 p. - ISBN 9780521682770
| Органоиди еукариотни клетки | ||
|---|---|---|
 |
||
Фондация Уикимедия. 2010 г.
- Железен хлорид
- Въглероден двуокис.
Вижте какво са „хлоропласти“ в други речници:
ХЛОРОПЛАСТИ- (от гръцки хлорос зелен и пластос оформен), вътреклетъчни органели (пластиди) на растенията, в които се извършва фотосинтеза; Благодарение на хлорофила те са оцветени в зелено. Намира се в различни клетки. тъкани от надземни растителни органи,... ... Биологичен енциклопедичен речник
ХЛОРОПЛАСТИ- (от гръцки хлорос зелен и пластос изваян образуван), вътреклетъчни органели на растителна клетка, в която протича фотосинтеза; оцветени в зелено (те съдържат хлорофил). Собствен генетичен апарат и... ... Голям енциклопедичен речник
Хлоропласти- тела, съдържащи се в растителните клетки, оцветени в зелено и съдържащи хлорофил. При висшите растения хлорофилите имат много определена форма и се наричат хлорофилни зърна; Водораслите имат разнообразна форма и се наричат хроматофори или... Енциклопедия на Брокхаус и Ефрон
Хлоропласти- (от гръцки хлорос зелен и пластос оформен, оформен), вътреклетъчни структури на растителна клетка, в която протича фотосинтеза. Те съдържат пигмента хлорофил, който ги оцветява в зелено. В клетката на висшите растения има от 10 до ... Илюстрован енциклопедичен речник
хлоропласти- (гр. хлорос зелен + формиращ lastes) зелени пластиди на растителна клетка, съдържащи хлорофил, каротин, ксантофил и участващи в процеса на фотосинтеза вж. хромопласти). Нов речник на чуждите думи. от EdwART, 2009. хлоропласти [гр.... ... Речник на чуждите думи на руския език
Хлоропласти- (от гръцки хлорос зелен и plastós оформен, образуван) вътреклетъчни органели на растителна клетка Пластиди, в които протича фотосинтеза. Те са оцветени в зелено поради наличието на основния пигмент на фотосинтезата... Велика съветска енциклопедия
хлоропласти- ов; мн. (единица хлоропласт, a; m.). [от гръцки chlōros бледозелен и пластос изваян] Botan. Тела в протоплазмата на растителните клетки, съдържащи хлорофил и участващи в процеса на фотосинтеза. Концентрация на хлорофил в хлоропластите. * * *… … енциклопедичен речник
Хлоропласти- тела, съдържащи се в растителните клетки, оцветени в зелено и съдържащи хлорофил. Във висшите растения X. имат много определена форма и се наричат хлорофилни зърна (виж); Водораслите имат различни форми и се наричат... ... Енциклопедичен речник F.A. Brockhaus и I.A. Ефрон
Хлоропласти- мн. Зелени пластиди на растителна клетка, съдържащи хлорофил, каротин и участващи в процеса на фотосинтеза. Обяснителен речник на Ефрем. Т. Ф. Ефремова. 2000... Съвременен тълковен речник на руския език от Ефремова
ХЛОРОПЛАСТИ- (от гръцки хлорос зелен и plastоs изваян, образуван), расте вътреклетъчни органели. клетки, в които протича фотосинтеза; оцветени в зелено (те съдържат хлорофил). Собствен генетични апаратура и протеинов синтез... ... Естествени науки. енциклопедичен речник
Хлоропластите са структури, в които протичат фотосинтетични процеси, водещи в крайна сметка до свързване на въглероден диоксид, освобождаване на кислород и синтез на захари. удължени структури с ширина 2-4 микрона и дължина 5-10 микрона. Зелените водорасли имат гигантски хлоропласти (хроматофори), достигащи дължина до 50 микрона.
зелените водорасли могат да имат по един хлоропласт на клетка. Обикновено има средно 10-30 хлоропласта на клетка от висшите растения. Има клетки с огромен брой хлоропласти. Например, около 1000 хлоропласта са открити в гигантските клетки на палисадната тъкан на бакланя.
Хлоропластите са структури, ограничени от две мембрани – вътрешна и външна. Външната мембрана, подобно на вътрешната, има дебелина около 7 микрона, те са разделени една от друга с междумембранно пространство от около 20-30 nm. Вътрешната мембрана на хлоропластите разделя пластидната строма, която е подобна на митохондриалната матрица. В стромата на зрелия хлоропласт на висшите растения се виждат два вида вътрешни мембрани. Това са мембрани, които образуват плоски, разширени стромални ламели и мембрани от тилакоиди, плоски дисковидни вакуоли или торбички.
Стромалните ламели (с дебелина около 20 µm) са плоски кухи торбички или имат вид на мрежа от разклонени и свързани помежду си канали, разположени в една и съща равнина. Обикновено стромалните ламели вътре в хлоропласта лежат успоредно една на друга и не образуват връзки една с друга.
В допълнение към стромалните мембрани, мембранните тилакоиди се намират в хлоропластите. Това са плоски, затворени мембранни торбички с форма на диск. Размерът на междумембранното им пространство също е около 20-30 nm. Тези тилакоиди образуват подобни на монети купчини, наречени грана. 
Броят на тилакоидите на грана варира значително: от няколко до 50 или повече. Размерът на такива стекове може да достигне 0,5 микрона, така че зърната се виждат в някои обекти в светлинен микроскоп. Броят на зърната в хлоропластите на висшите растения може да достигне 40-60. Тилакоидите в граната са близо един до друг, така че външните слоеве на техните мембрани са тясно свързани; на кръстовището на тилакоидните мембрани се образува плътен слой с дебелина около 2 nm. В допълнение към затворените камери на тилакоидите, граната обикновено включва и участъци от ламели, които също образуват плътни 2-nm слоеве в точките на контакт на техните мембрани с тилакоидните мембрани. По този начин стромалните ламели изглежда свързват отделните грани на хлоропласта един с друг. Въпреки това, кухините на тилакоидните камери винаги са затворени и не преминават в камерите на междумембранното пространство на стромалните ламели. Стромалните ламели и тилакоидните мембрани се образуват чрез отделяне от вътрешната мембрана по време на началните етапи на развитие на пластид.
ДНК молекулите и рибозомите се намират в матрицата (стромата) на хлоропластите; Тук става и първичното отлагане на резервния полизахарид, нишестето, под формата на нишестени зърна.
Характерна особеност на хлоропластите е наличието на пигменти, хлорофили, които придават цвета на зелените растения. С помощта на хлорофила зелените растения абсорбират енергия от слънчевата светлина и я превръщат в химическа енергия. 

Функции на хлоропластите
Пластиден геном
Подобно на митохондриите, хлоропластите имат собствена генетична система, която осигурява синтеза на редица протеини в самите пластиди. ДНК, различни РНК и рибозоми се намират в матрицата на хлоропласта. Оказа се, че ДНК на хлоропластите рязко се различава от ДНК на ядрото. Представен е от циклични молекули с дължина до 40-60 микрона, с молекулно тегло 0,8-1,3x108 далтона. В един хлоропласт може да има много копия на ДНК. Така в индивидуален царевичен хлоропласт има 20-40 копия на ДНК молекули. Продължителността на цикъла и скоростта на репликация на ядрената и хлоропластната ДНК, както е показано в клетките на зелените водорасли, не съвпадат. Хлоропластната ДНК не е в комплекс с хистони. Всички тези характеристики на хлоропластната ДНК са близки до характеристиките на ДНК на прокариотните клетки. Освен това, сходството на ДНК на хлоропластите и бактериите е допълнително подсилено от факта, че основните транскрипционни регулаторни последователности (промотори, терминатори) са еднакви. Всички видове РНК (информационна, трансферна, рибозомна) се синтезират върху хлоропластна ДНК. Хлоропластната ДНК кодира рРНК, която е част от рибозомите на тези пластиди, които принадлежат към прокариотния тип 70S (съдържат 16S и 23S рРНК). Хлоропластните рибозоми са чувствителни към антибиотика хлорамфеникол, който инхибира протеиновия синтез в прокариотните клетки.
Точно както в случая с хлоропластите, отново се сблъскваме със съществуването на специална система за синтез на протеини, различна от тази в клетката.
Тези открития подновиха интереса към теорията за симбиотичния произход на хлоропластите. Идеята, че хлоропластите са възникнали чрез комбиниране на хетеротрофни клетки с прокариотни синьо-зелени водорасли, изразена в началото на 19-ти и 20-ти век. (А.С. Фомицин, К.С. Мережковски) отново намира своето потвърждение. Тази теория се подкрепя от удивителното сходство в структурата на хлоропластите и синьо-зелените водорасли, сходството с основните им функционални характеристики и най-вече със способността за фотосинтетични процеси.
Има много известни факти за истинска ендосимбиоза на синьо-зелени водорасли с клетки от низши растения и протозои, където те функционират и снабдяват клетката гостоприемник с фотосинтетични продукти. Оказа се, че изолираните хлоропласти също могат да бъдат избрани от някои клетки и използвани от тях като ендосимбионти. При много безгръбначни (ротифери, мекотели), които се хранят с висши водорасли, които усвояват, непокътнатите хлоропласти попадат в клетките на храносмилателните жлези. Така при някои тревопасни мекотели в клетките са открити непокътнати хлоропласти с функциониращи фотосинтетични системи, чиято активност се наблюдава чрез включването на C14O2.
Както се оказа, хлоропластите могат да бъдат въведени в цитоплазмата на миши фибробластни култури чрез пиноцитоза. Те обаче не са били атакувани от хидролази. Такива клетки, които включват зелени хлоропласти, могат да се делят в продължение на пет поколения, докато хлоропластите остават непокътнати и извършват фотосинтетични реакции. Правени са опити за култивиране на хлоропласти в изкуствена среда: хлоропластите могат да фотосинтезират, в тях се извършва синтез на РНК, те остават непокътнати в продължение на 100 часа, а деленията се наблюдават дори в рамките на 24 часа. Но след това имаше спад в активността на хлоропластите и те умряха.
Тези наблюдения и редица биохимични работи показаха, че тези характеристики на автономност, които притежават хлоропластите, все още са недостатъчни за дългосрочно поддържане на техните функции, още по-малко за тяхното възпроизводство.
Наскоро беше възможно напълно да се дешифрира цялата последователност от нуклеотиди в цикличната ДНК молекула на хлоропластите на висшите растения. Тази ДНК може да кодира до 120 гена, сред които: гени на 4 рибозомни РНК, 20 рибозомни протеини на хлоропласти, гени на някои субединици на хлоропластна РНК полимераза, няколко протеини на фотосистеми I и II, 9 от 12 субединици на АТФ синтетаза, части на протеини от комплексите на електронната транспортна верига, една от субединиците на рибулоза дифосфат карбоксилаза (ключовият ензим за свързване на CO2), 30 tRNA молекули и още 40 все още неизвестни протеина. Интересното е, че подобен набор от гени в хлоропластната ДНК е открит в такива далечни представители на висши растения като тютюн и чернодробен мъх.
По-голямата част от хлоропластните протеини се контролира от ядрения геном. Оказа се, че редица от най-важните протеини, ензими и съответно метаболитните процеси на хлоропластите са под генетичен контрол на ядрото. По този начин клетъчното ядро контролира отделните етапи на синтеза на хлорофил, каротеноиди, липиди и нишесте. Много ензими от тъмен стадий и други ензими, включително някои компоненти на електронната транспортна верига, са под ядрен контрол. Ядрените гени кодират ДНК полимераза и аминоацил-тРНК синтетаза на хлоропласти. Повечето рибозомни протеини са под контрола на ядрени гени. Всички тези данни ни карат да говорим за хлоропластите, както и за митохондриите, като структури с ограничена автономност.
Транспортът на протеини от цитоплазмата до пластидите се извършва по принцип подобен на този на митохондриите. Тук също в точките на конвергенция на външната и вътрешната мембрана на хлоропласта се намират каналообразуващи интегрални протеини, които разпознават сигналните последователности на синтезираните в цитоплазмата хлоропластни протеини и ги транспортират до матрицата-строма. От стромата внесените протеини, според допълнителни сигнални последователности, могат да бъдат включени в пластидните мембрани (тилакоиди, стромални ламели, външни и вътрешни мембрани) или локализирани в стромата, като част от рибозоми, ензимни комплекси от цикъла на Калвин и др.
Удивителното сходство на структурата и енергийните процеси в бактериите и митохондриите, от една страна, и в синьо-зелените водорасли и хлоропластите, от друга, служи като силен аргумент в полза на теорията за симбиотичния произход на тези органели. Според тази теория появата на еукариотна клетка преминава през няколко етапа на симбиоза с други клетки. На първия етап клетки като анаеробни хетеротрофни бактерии включват аеробни бактерии, които се превръщат в митохондрии. Успоредно с това в клетката гостоприемник прокариотният генофор се образува в ядро, изолирано от цитоплазмата. Ето как могат да възникнат хетеротрофни еукариотни клетки. Повтарящите се ендосимбиотични връзки между първичните еукариотни клетки и синьо-зелените водорасли доведоха до появата на структури от хлоропластен тип в тях, което позволява на клетките да извършват автосинтетични процеси и да не зависят от наличието на органични субстрати (фиг. 236). По време на формирането на такава съставна жива система, част от генетичната информация на митохондриите и пластидите може да се промени и да бъде прехвърлена в ядрото. Например, две трети от 60-те рибозомни протеини на хлоропластите са кодирани в ядрото и синтезирани в цитоплазмата и след това интегрирани в рибозомите на хлоропластите, които имат всички свойства на прокариотните рибозоми. Това движение на голяма част от прокариотните гени в ядрото доведе до факта, че тези клетъчни органели, запазвайки част от предишната си автономия, попаднаха под контрола на клетъчното ядро, което до голяма степен определя всички основни клетъчни функции.
Пропластиди
При нормално осветление пропластидите се превръщат в хлоропласти. Първо, те растат, с образуването на надлъжно разположени мембранни гънки от вътрешната мембрана. Някои от тях се простират по цялата дължина на пластида и образуват стромални ламели; други образуват тилакоидни ламели, които се подреждат, за да образуват граната на зрелите хлоропласти. Развитието на пластидите протича малко по-различно на тъмно. При етиолираните разсади обемът на пластидите, етиопластите, първоначално се увеличава, но системата от вътрешни мембрани не изгражда ламеларни структури, а образува маса от малки везикули, които се натрупват в отделни зони и дори могат да образуват сложни решетъчни структури (проламеларни тела). Мембраните на етиопластите съдържат протохлорофил, жълт предшественик на хлорофила. Под въздействието на светлината от етиопластите се образуват хлоропласти, протохлорофилът се превръща в хлорофил, синтезират се нови мембрани, фотосинтетични ензими и компоненти на електронтранспортната верига.
Когато клетките са осветени, мембранните везикули и тръбички бързо се реорганизират и от тях се развива цялостна система от ламели и тилакоиди, характерни за нормален хлоропласт.
Левкопластите се различават от хлоропластите по липсата на развита ламеларна система (фиг. 226 b). Те се намират в клетките на складовите тъкани. Поради неопределената си морфология, левкопластите трудно се различават от пропластидите, а понякога и от митохондриите. Те, подобно на пропластидите, са бедни на ламели, но въпреки това са способни да образуват нормални тилакоидни структури под въздействието на светлина и да придобиват зелен цвят. На тъмно левкопластите могат да натрупват различни резервни вещества в проламеларните тела, а зърната от вторично нишесте се отлагат в стромата на левкопластите. Ако така нареченото преходно нишесте се отлага в хлоропластите, което присъства тук само по време на асимилацията на CO2, тогава в левкопластите може да се получи истинско съхранение на нишесте. В някои тъкани (ендосперма на зърнени култури, коренища и грудки) натрупването на нишесте в левкопласти води до образуването на амилопласти, напълно пълни с резервни нишестени гранули, разположени в стромата на пластида (фиг. 226c).
Друга форма на пластид във висшите растения е хромопластът, който обикновено пожълтява в резултат на натрупването на каротеноиди в него (фиг. 226d). Хромопластите се образуват от хлоропласти и много по-рядко от техните левкопласти (например в корените на морковите). Процесът на избелване и промени в хлоропластите се наблюдава лесно по време на развитието на венчелистчетата или по време на узряването на плодовете. В този случай в пластидите могат да се натрупат жълти капчици (глобули) или в тях да се появят тела под формата на кристали. Тези процеси са свързани с постепенно намаляване на броя на мембраните в пластида, с изчезването на хлорофила и нишестето. Процесът на образуване на цветни глобули се обяснява с факта, че при разрушаването на ламелите на хлоропластите се отделят липидни капчици, в които различни пигменти (например каротеноиди) са добре разтворени. По този начин хромопластите са дегенериращи форми на пластиди, подложени на липофанероза - разпадането на липопротеиновите комплекси.